この書類はネイチャーに提出した二報論文に対する査読書です。最初がアーティクル論文に対するもの、もう一つがレター論文に対するリヴァイズ要請です。最初の査読対象になったオリジナルはありませんので、図表等は現在のリトラクトされた最終論文とは違うので推測していくしかない。

まず題名はオリジナルでは<Stimulaus-Triggered Fate Conversion of Somatic Cells into Pluripotency>であった。最終稿のタイトルも同じですが大小文字の使い分けを変えていますね。<Stimulus-triggered fate conversion of somatic cells into pluripotency>となっている。何故かは分かりません。
レフェリーのコメントはそもそも専門的なことなのでど素人には分かりにくいのですが、何よりもオリジナルの論文の文面と図が無いので何を言わんとしているのかの推測がとても難しい。専門であっても結構むつかしいのではないかと思いますが、門外ですから常識の範囲にないことの推測は大変です。
パラグラフごとに翻訳していきましょう。
>レフェリー・コメント
>
>リフェリー #1 (著者への所見)
>
>この現在の論文では、著者らは、多能性へ向けた細胞核のリプログラム過程での細胞への刺激の役割を重要視している。これはとても興味深く、恐らく革新的な論文である。しかし、結論に導く説明とデータは幾分推論に過ぎず、かつ、一部は手始めの段階である。
>
ちなみにこの#1は2011年に三誌論文の最初に投稿したネイチャーの査読者と同じ人だと桃子が言ってますね。桃子本308P。
>>
まず目を引いたのは、ネイチャー編集者のコメントの変化だった。
初稿当時には「論文は掲載できない」と淡々とした文面で伝えたのに対し、再投稿時には大きな関心を示し、六カ月以内に追加実験をしたうえで、改訂するよう強く勧めて、「改訂版の原稿をぜひ見たい」と記した。これを読んだある大学教授は「熱狂的といってもいいような反応、初稿当時とあまりに違う」と驚いた。
一方、査読者たちは、「発生生物学や幹細胞生物学の転換を迫る発見」「特筆すべき発見」などと賞賛しつつも、問題点や疑問点を多数挙げていた。
人数は一人増えて三人になっていたが、一人目の査読者が初投稿時の一人目と同じ人物であることは明らかだった。アーティクル論文に関するコメントの最初の一文が全く同じな上に、同様のフレーズが何度も登場したからだ。初投稿時は前向きな表現が見当たらなかったが、再投稿時は「非常に興味深く、画期的な可能性を秘めている」と評価する言葉があり、「結論を支えるデータと説明は非常に推論的で予備的だ」という一文は「結論を支えるデータと説明は少々推論的で、いくつかのケースでは予備的だ」と弱められていた。
おそらく同じメンバーがレター論文も査読しており、一人目の査読者はレターに関するコメントの最後にこう指摘した。-----著者達はSTAP細胞が次世代の再生医療の材料として有望だと主張するが、両論文中にはES細胞やiPS細胞と比較してSTAP細胞の(材料としての)品質を評価する実験が一つもない。ゲノムレベルでの詳細な解析をしない限り、この論文の掲載を勧められない。私は現在のデータを疑いはしないが、プロセスを要約すれば「魔法のような(magical)」アプローチだ-----。
編集からのメッセージ中で中央に大きく墨消しのある部分に桃子の書いている<六カ月以内に追加実験をしたうえで、改訂するよう強く勧めて、「改訂版の原稿をぜひ見たい」と記した。>内容があったのでしょうね。なぜこの部分が墨で消されているのかも流出元の配慮を推測する上での疑義の一つです。桃子が見た時にはこの文面は消されていないのですから、
①桃子が消したか、
②私がそのホームページからの入手した慶応大学の吉村氏が消したか、
③吉村氏がこんな流出してはいけない筈の査読書がネットに流されていると紹介したもとの書類に墨消しが既にあったのか、
のいずれかです。桃子は本に書いてますから①はない。②は意図的に吉村氏が流したのなら、ネイチャー誌の積極姿勢は隠してあげようと配慮したことになる。③の場合は、桃子は拡散してはいけない多くの入手書類を意見を聞くという名目で何人かの学者に配布している。その中の誰かが意図的に流したのならこの部分を隠す理由は弱いんですね。言うまでもなく吉村氏は遠藤論文の謝辞の中に出て来る仲間ですから、どうも②の香りがしますかね。
この第一レフェリーが誰であるかに関しては全世界的にもとても狭い社会なので仲間内では推測可能なのではないでしょうかね。三誌論文がすべてリジェクトされた時に若山さんは以下のように言ったとされている。手記103P。
>>
春から投稿されていたスフェアの論文は、残る最後の三大誌であるサイエンスからも、レビューワーにはまわるものの2011年8月21日には不採択の連絡が届けられた。若山先生から、「一度不採択になった論文は通常もう一度投稿することができないが、幹細胞株化の論文と2報同時に投稿すれば、再考されると思う。三大誌3つのレビューワーコメントの中で、ネイチャーからのコメントが一番やさしいので、ネイチャーに現在投稿を試みている論文と幹細胞株化の論文を2報同時投稿しよう」と提案され、幹細胞株化の論文の執筆とともに現在投稿中の論文に対するネイチャーからのレビューワーコメントへの反論を用意するように指示された。また、ES細胞の混入による可能性を否定するために、「ES細胞は同時に培養していなかった」という記事を原稿に加えることも指示された。
最初のネイチャー誌投稿は2012年の4月ですから投稿論文の実験中に培養されているES細胞は小保方さんの持っている学生のES細胞以外は無いのですが、4月末から5月初旬にFLSのジャームライントランスミッション結果が出ていて、サイエンスには二報同時論文と同じ結果図が載せられている筈ですからそこにコントロールESキメラのジャームライントランスミッション結果も載せられていたはずです。小保方さんはキメラを作れませんから以下のデータのESキメラも若山さんが実験している筈のものです。

サイエンスは8月21日にリジェクトされたことは分かっていますが何時投稿したのかを理研は発表していません。全部分かっている筈ですね。桃子は全部の書類を入手している。20日前だとしてみましょう。
8月1日だとしてみるのです。ESのデータは当然その時には出ているわけです。まず生きて生まれた数が書かれていて、成長できた数も書かれている。adulthoodというのは生後何日までを見るのかは知りませんので、生殖能力を得るのは生後50日後とされていますが、ここでは10日は生きて生育していたという意味にとっておきましょう。この場合生まれたのは7月20日前後ということになる。この前に生まれて来るのに20日掛かります。交配は7月1日前後です。交配できるようになるのに親のキメラマウスは50日掛かります。逆残すると親のESキメラマウスが生まれたのは5月10日前後になる。この時点で既に若山さんの事後MTA の細胞リストに記載されたコントロールES細胞の樹立日が5月25日になっているのは嘘だということになってしまうんですね。これはadulthood期間の10日仮定をゼロ日にしても間に合っていませんね。更に、キメラを作るのにインジェクト後20日かかる。インジェクションは4月20日前後だということになるのです。
若山さんの事後MTAの細胞リストは2014年4月1日付で提出されている。ここにはコントロールES細胞の樹立は2012年5月25日とされている。しかし、その後、桂報告書が2014/12/25に公開された時、その細胞リストには2012/4/19の樹立開始と書かれていたのです。本文中には以下のようにある。
>>
4)他の細胞株における遺伝的不均一性 この遺伝的不均一性は、129 CAG-GFP マウスに由来する他の細胞株にも反映していた。 若山氏により 129B6F1CAG-GFP マウスの独立した胚より複数の受精卵 ES 細胞株が樹立 されているが(129B6 F1ES1~6、2012 年 5 月作製)、いずれも第 6 染色体中程に B6 ホモ 領域を有していた。
細胞リストの樹立日は培養開始日と注されているが、事後MTA細胞リストの5月25日なら35日継代しているので11回も継代していることになる。増殖率実験をしているのではありませんから3継代もしたら樹立としてキメラ実験に使用するのが普通ではないか。しかも桂報告書はリストでは4月19日培養開始として本文中では5月作製としか書いてない。これだと5月1日であった事にもなりうる。意図的にごまかしているんですね。明確に共犯だということが分かりますね。事後MTAは木星さんが取り寄せて分かったので、これが無かったら国民は騙されたままであったでしょうね。
コントロールのES細胞キメラのジャームライントランスミッション実験は実際には行われていないと思いますね。若山さんのデータ捏造です。それを何とかごまかそうとして桂報告書は細胞リストに4月19日培養開始という嘘のデータを書き込んだ。それでもまだ間に合ってはいませんが、他方で4月はネイチャー誌への投稿(投稿日不明)とヴァカンティ氏の米国特許仮申請(4/24)があった。
最初のネイチャー論文にはジャームライントランスミッション実験は終わっていませんから書き込まれていません。そもそも小保方酸浴細胞核使用ntESの生殖能力確認ですから、リクルートのために書かせている論文とは無関係だったんです。4月のネイチャーリジェクトで後はヴァカンティ氏が自分の主催誌であるティシュー誌に掲載し、小保方さんを解放して自分に渡してくれるという話になってた筈だったわけです。ところがヴァカンティ氏がセルだサイエンスだと言い始めたのです。
ですから若山さんがネイチャーへの再提出に向けて<ES細胞の混入による可能性を否定するために、「ES細胞は同時に培養していなかった」という記事を原稿に加えることも指示>したのは別に嘘ではないわけです。まだ4月の論文提出時には129B6F1ES1~6は存在してなかった。小保方さんにそう言ってますから、逆に4/19より前にはこの作成時期を遡らせることが出来なかったんでしょうね。これは前提通りに作製していくと、キメラ胚へのインジェクションが4月20日前後ならその時にはES細胞は出来ていることになって、ES細胞は5日胚のインナーセルマスを取り出して培養を3継代程度確認したものでしょうからF1マウスの交配から12日程度かかっている。4月8日には計画しているということになる。私はESのジャームライントランスミッションなんて分かり切っていることなので若山さんがこんな実験をするわけがないと思いますね。予算の無駄です。ES細胞の増殖率実験も同じですね。やるわけがない。小保方さんにやらせるわけもないということです。対して小保方酸浴細胞核使用ntES細胞に関してはすべて確認しないといけません。未経験の細胞です。
第一レフェリーを誰それだと悟って若山さんがコンタクトをとっていたという可能性も考えられるんですが今はその問題は置いておきましょう。結構ややこしい。
それよりもこのレフェリーは刺激惹起の論旨を頭から否定しているので桃子が論調が変わったと言っているのは、二度目の提出で、しかも理研と笹井さんの名がありますから世辞も述べますね。前回のはヴァカンティ氏の名前です。
内容は厳しいものですね。これを若山さんが<三大誌3つのレビューワーコメントの中で、ネイチャーからのコメントが一番やさしいので>と言うところに何か第一レフェリーとの関係を疑義するわけです。
続きです。

>
>研究と結論は意図的にある特定の分野に焦点が当てられていて、他は除かれている。本文から読者は直ぐに刺激が多能性に向けた何らかのリプログラミングを引き起こしていると思ってしまう。このことはテラトーマとキメラ形成によって際立って提示されている。しかし、リプログラミングは一つの説明であるにすぎない。再生、脱分化及び増殖能力、そして異なる組織への分化は又腫瘍を含む癌幹細胞の特徴でもある。私は著者らに遺伝子操作なしの幹細胞様の作成法の発見に関する主張に注意深くあることをお奨めしたい。著者らは処置が遺伝子に引き起こしている実際の効果を調べ、遺伝子の不安定性に気づかないといけない。
論旨の全否定だということはアーティクル論文を読んでいる人には自明ですね。刺激惹起というのは手記では笹井さんが命名したと花を持たせた書き方になってますが、SACsという言葉は例の実験ノートにも以下のように出ている。
無論サイエンス誌投稿論文ではStress Altered Cellsという命名で論じられていて、刺激惹起という発想そのものはもともと小保方さんが抱いていた仮説ですが、これを第一レフェリーはアーティクル論文においても全否定しているということです。
本文です。
>
>著者らは、体細胞系譜決定が遺伝子修飾に働きかけることによってのみ強力に変化させ得ると、又、その過程は自然には生じないと、読者を信じるように導いている。遺憾ながら私は同意しない。これは心筋細胞の部分的分化が心臓疾患治癒の原因となって以降のケースとは違う(Senyo et al.,2012 nature 事例を参照)。面白いことに、細胞系譜決定の動的平衡が癌幹細胞培養維持にも又示されてきている(Gupte et al.,2011 Cell 事例)。私見では、これらの事例は、著者らによって為されている主張に反して、哺乳動物体細胞は遺伝子修飾無しに、脱分化能力を持つことを示している。
ここはど素人としてはどちらが正しいのかを判断することはできませんね。笹井さんはただ英語と論文の書き方の指導をしてやって、この論文を通してやってくれと頼まれている。個々の実験に虚偽があるかもしれないと考えては出来ない仕事を頼まれている。一科学者としてとか、CDBのGD、つまり管理職としてどうあるべきかという判断は結構むつかしい判断で、笹井さんほどの人であるとこの論文はかなり実験の詰めが甘いということはすぐに感じたと思いますが、ただし、キメラがスタンダードな意味で書かれている通りに出来たということは疑えません。そもそも論文を通してやってくれと頼まれたことそのこともキメラができているという絶対的事実に基づているわけですから、ここを疑えということは、若山さんを疑うということであり、理研のシニアな学者を疑うということですから、常識的にまずは不可能です。そういう人なら理研に10年間も居た事自体の管理責任がどうなのだということになる。10年間そんなことは露なかったから10年満期勤務したので何かいけないかと言われても非難のしようがないのではないでしようか。
では、小保方さんが不正をしてキメラが出来たのだとして、この場合関係者全員が小保方さんに騙されたということになるわけですが、そもそも若山さんがキメラが出来たと証言しているのだから、若山さんが小保方さんに騙されて、キメラを作ったのだとしたら、関係者全員が騙された責任は若山さんにあるわけです。これを聞いて面白いと思った西川さんや、竹市さんや、笹井さんや、丹羽さんや、野依さんが信じ込んだのは若山さんが出来たと言ったからで、西川さんや、竹市さんや、笹井さんや、丹羽さんや、野依さんが信じ込んだのを間違いだということはできないということをまず確認しておかなけければなりませんよね。
若山さんを騙した小保方さんが悪いという話はその後に論ずべきことですから、小保方さんが若山さんを騙したから、西川さんや、竹市さんや、笹井さんや、丹羽さんや、野依さんが騙されたというのは間違いですね。混同しないようにしないといけませんよね。それとこれは違いますね。
まずはすべては若山さんが悪いのです。
次に小保方さんが若山さんを騙したのが悪いというのも又おかしな話で、若山さんは小保方さんに騙されてはいけない人です。武田邦彦教授は最初にここを指摘していますよね。「僕は自分の学問分野で自分の学生に騙されたりはしないよ。騙されないくらいの圧倒的知識差があるから教授をやってる。」と。これを事件の話に置き換えると、「若山さんは自分の学問分野で小保方さんに騙されたりはしないよ。騙されないくらいの圧倒的知識差があるから教授をやってる。」
若山さんは山梨大の教授として、そこに小保方さんを助手で連れて行こうとしたのです。
武田教授の説明が正しいなら、若山さんは小保方さんに騙されたことなど一度もないはずですよね。むしろ上から見ていたはずですし、こんなことは別に学問分野でなくてもどんな職場でもそうですね。普通は上から下はミエミエですよね。それが普通でないのだったら、人間は成長しないものだということになってしまう。
騙したのなら若山さんが小保方さんを騙したということしか考えられませんよね。それが世間常識というものです。それに何か目下が悪いことしたらどやしつけて内々で処理して外に恥をさらすなんてことはしませんよね。ところが若山さんは内輪で話し合うこともせずに「騙されたあ」と世間に公言しているんです。おかしいと思わない一般人はいません。
従って、笹井さんが学問的にこの論文をおかしいと思わなかったことを非難することはできません。仮に笹井さんが第一査読者であったら同じことをコメントしたかもしれませんよね。なぜなら第一レフェリーはキメラが出来たという若山さんの言葉を信じなければならないような義務もなければ、若山さんと同じ職場でしかもGDとして管理者の立場にあった関係を若山さんとの間に持っているわけでもないからです。笹井さんだって、同じく関係ない立場だったらもっと厳しいコメントすらしていても不思議はないでしょうね。
でも笹井さんは同じ研究所に属している研究仲間です。その若山さんが例えば何か、嘘でも、とても馬が合わずに嫌いな人であったとしてすら、キメラが出来たと言っていることを疑うということはあり得ないことで、そもそもそんな考えではこの研究の仕事はできません。第一レフェリーでもこの論文を自分の仲間が書いていたらキメラができていたことは信じざるを得ないのですから、論文を信じるということになる。信じないときにはだから何かアーティファクトが無いかと訝ることになるわけです。
従って三誌論文とも皆が皆ESコンタミではないかと疑ったのです。ここでもレフェリーはコンタミを疑っている。しかし、真実はESコンタミキメラではなくて小保方酸浴細胞核使用ntESキメラだっただけですね。若山さんの最初の他愛ない嘘が最後の最後まで明かされないままになってしまっただけの事件でしたね。若山さんは最後まで論文がリジェクトされると信じていたんです。それまではすべてうまくリジェクトされてきていますからね。
若山さんが山梨大に転居して後の二報同時を主張しているのは、実は裏で、ネイチャーのこの第一レフェリーに本当のことを言ってないかとも疑義していますが、今は考察がややこしくなるので置いているところですね。
酸浴させて癌化している細胞は、エピジェネティクな遺伝子修飾がリプログラムされていなくても、<遺伝子修飾無しに、脱分化能力を持つ>と言われても、笹井さんはキメラができていると信じてますからね。この第一レフェリーは出来てないと思ってるんです。できていたと思った瞬間に<遺伝子修飾無しに、脱分化能力を持つ>なんて主張は関係なくなる。細胞の細かい解析がどうであれ、キメラができていたら多能性細胞であることは証明され終わっている。遺伝子修飾されたままにキメラができるなんてことはありませんからね。
本文です。
>
>施された細胞処置が遺伝子異常を引き起こす可能性に加えて、大量細胞死を引き起こしていることも明白である。著者らはその後の実験の流れに沿って、生き残った細胞に意識を向けて、図1に細胞数計測見積を示しているが、それは恐らく最善の実験ではない。著者らは細胞死の数量を実証し、かつ示し、そしてこれをOct4-GFP観測と同時にCD45陽性細胞を共染色することで実行しなければならない。これはPI/Annexin-Vを使ってできる簡単な実験である。
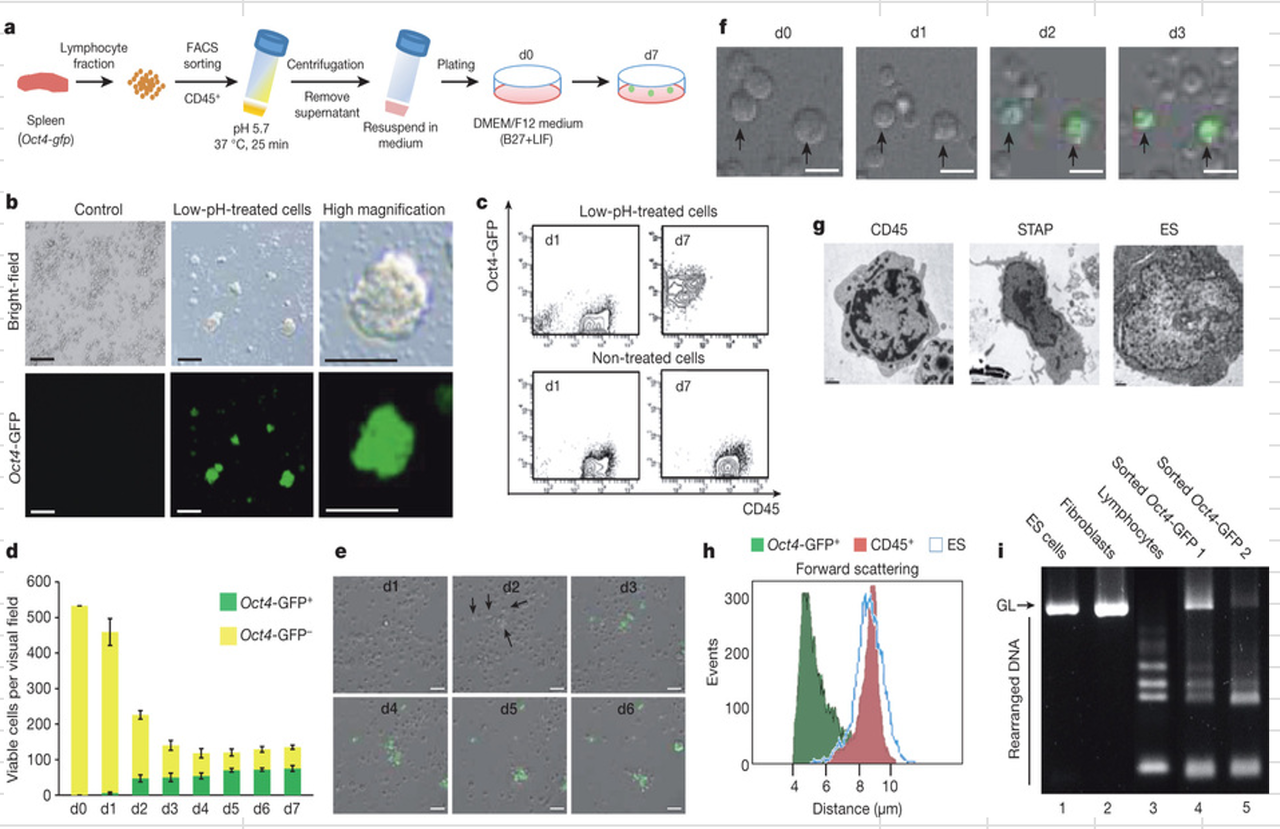
この図1はオリジナルでどうなっていたのかは分かりませんが、最終版の図のFigure1-dに結果しているのではないでしょうかね。

レジェンドは以下です。
d, GFP+ (green) and GFP− (yellow) cell populations (average cell numbers per visual field; ×10 objective lens). n = 25; error bars show average ± s.d.
d,GFP +(緑)、GFP-(黄色)の細胞集団(視野当たりの平均細胞数、×10対物レンズ)。 試行数= 25;エラーバーは、平均±のs.d.を表示。
PI/Annexin-Vというのは死細胞を染色してFACS選別する実験のようですが、これはアーティクル論文の海賊版翻訳を中断させていますが、再開すべきですね。
まずFACSの実験は以下の二つです。
Figure 1-c

レジェンドは以下です。
c, FACS analysis. The x axis shows CD45 epifluorescence level; y axis shows Oct4-GFP level. Non-treated, cultured in the same medium but not treated with low pH.
c,FACS解析。 x軸はCD45の落射蛍光レベルを示す:y軸はOct4-GFPのレベルを示す。非処理(細胞)は同一培地であるが、低pH処理していない。
Extended Data Figure 1-b

レジェンドは以下です。
b, Gating strategy for Oct4-GFP+ cell sorting. Top: representative results 7 days after the stress treatment. Bottom: non-treated control. P3 populations were sorted and counted as Oct4-GFP+ cells for all experiments.
b、Oct4-GFP陽性細胞ソーティングのゲーティング計画。上:ストレス治療から7日後の代表的な結果。下:未処理のコントロール。 P3集団を分類し、すべての実験でOct4-GFP陽性細胞としてカウントした。
そしてPI/Annexin-Vの実験は以下のExtended Data Figure 1-g,hです。

レジェンドは以下です。
g, h, Time course of apoptosis after the low-pH exposure. Stress-treated cells and non-stress-treated control cells were stained with CD45, annexin-V and propidium iodide at day 0 (immediately after stress treatment), day 3 and day 7. g, Blue bars, GFP+CD45−; orange bars, GFP−CD45+. Percentages in total cells included propidium-iodide-positive cells. h, Annexin-V-positive cells in these cell populations were analysed by FACS.
g、h、低pH曝露後のアポトーシスの時間経過。ストレス処理細胞および非コントロールの非ストレス処理細胞は、0日目(ストレス処理直後)、3日目および7日目に、CD45、アネキシンVおよびヨウ化プロピジウムで染色された。g、青色バー、GFP陽性CD45陰性。オレンジ色バー、GFP陰性CD45陽性。全細胞のパーセンテージにはヨウ化プロピジウム陽性細胞が含まれている。 h、これらの細胞集団におけるアネキシンV陽性細胞をFACSによって分析した。
まずFigure 1-cに関しては<海賊版翻訳アーティクル1>で以下のように考察した。
>>
**********
FACS( Fluorescence activated cell sorter)は蛍光活性化セルソーティングと訳されている。細胞にレーザー光を当てて識別後、電荷を与えて弾き飛ばす原理で分類できるらしい。
蛍光顕微鏡(fluorescence microscope)は強い励起光と弱い蛍光をフィルターを使って分離しているが、epifluorescence microscope というのがあって落射蛍光顕微鏡と呼ばれているらしいが、励起光を顕微鏡の中を通して照射するシステムで、ミラーをうまく使って蛍光のみを見たり撮影したりできる装置のようです。これはこれで専門的な知識が必要なようで、小保方さんは使い方を間違ってるとか言われていたりもしているが、ど素人には判断がつきませんね。
ただ、この画像はそんなに難しい操作結果だとも思えませんけどね。CD45+細胞は7日間変化がなく、だいたい同じ場所に飛ばされて集まっているが、酸浴細胞は1日目はたんなるCD45+細胞なのに7日目の細胞はGFP識別でほとんどが別の位置に飛ばされているということでしょう。
検出器の前を通るごとに識別されて分類される器械のようですね。こういう物理工学分野は素人にもすごいと分かりやすいが細胞はへなへなとしていて分かりませんねえ。
epifluorescence levelという意味は、光度指標か何かがあるのかどうかは分かりません。CD45+は何か抗体で蛍光染色しているはずですし、Oct4-GFPは発現すれば励起光を当てて確認するのでしょうが、両軸共にその光度か何かの目盛りにどんな意味があるのかは分かりませんね。10の何乗かという表示らしい目盛りの数値もよく見えませんが、ルクス表示のようですね。照度のようですね。
丹羽さんのGFPの漏れ出しは最大10%でした。この図はほぼすべてGFP発現していることになってますよね。蛍光しているスフィア塊だけを分析してますよね。因みに、言うまでもありませんが自家蛍光細胞はGFPでは選別されませんよね。自家蛍光だなんて言ってた人たちのお里の知れるところです。腹に一物あるんですね。ただ、その一物の道徳的評価はまだそんな判断の出来る段階ではありませんね。
**********
結論的には酸浴未処理細胞と酸浴細胞の酸浴直後のDAY0ではすべてがCD45陽性細胞(白血球)であるのに対して、酸浴DAY7では全部がOct4-GFP蛍光細胞になっているということです。
ただ、これは素人なので分からないのですが、これはCD45陽性でかつGFP陽性であった時には理論的には対角線上の辺りに集まると思いますが、全体がCD45の発現が弱まって左に寄っている。それが白血球が多能性細胞にリプログラムされたからなのか、それとも酸浴で白血球表面のCD45抗体蛋白が壊れて、その代わりに丹羽さんの発見したGFPの漏れ出しを検知しているのかは研究しないと分かりませんよね。
しかし再現実験ではキメラは出来ませんでしたので、この細胞がES並みの多能性細胞であることはないのですから、何かのアーティファクトなんですよね。ですから、ES細胞で若山さんにキメラを作らせたような人がこんなに真面目に研究しているわけがないと知れるところです。
論文ではキメラは出来ていると信じていますから、この現象を正しく考え直すなんてことは出来ませんよね。丹羽さんの発見したGFPの漏れ出しに気づくためには、キメラができないという事実から出発しないと無理なんです。小保方さんも、笹井さんもキメラは出来ているという前提でこの現象を見ている。
キメラができているという前提ならこのGFP発現は多能性細胞への転換だと考える方が当たり前だということに気づかないといけないですよね。ですから、査読者の「遺憾ながら私は同意しない」という疑義は必ずキメラは出来てないという根本的疑義に基づくことになるのです。
Extended Data Figure 1-bも<海賊版翻訳アーティクル1>で考察していますので省略しますが、何しろど素人なので機械の原理に関して無知ですから誤解も多いでしょうね。
Figure1-dはまさに査読者が「著者らはその後の実験の流れに沿って、生き残った細胞に意識を向けて、図1に細胞数計測見積を示している」と言っていて、かつその前に「研究と結論は意図的にある特定の分野に焦点が当てられていて、他は除かれている。本文から読者は直ぐに刺激が多能性に向けた何らかのリプログラミングを引き起こしていると思ってしまう。このことはテラトーマとキメラ形成によって際立って提示されている。しかし、リプログラミングは一つの説明であるにすぎない。」というアーティクル論文の論旨全体に対する全否定の向けられたまさにその解釈ですね。
笹井・小保方論文ではこの緑の細胞がキメラになったのだと解釈されている。実際キメラになったんです。論文はキメラになったという前提で書かれているんです。リヴァイズ実験中に理研内部でキメラになってないなどという解釈はありませんから、査読者はその解釈は可能性の一つに過ぎなくて他の解釈もありうると批判しているわけですが、笹井・小保方さんにはそんな視点は無い。
査読者はキメラはESコンタミだと思っていますから、別の解釈もありうることになるわけですが、この査読書は採用を考えているので、リヴァイズしてくれと要請してきているものですから、当然キメラが出来たことを認めているんですね。三誌論文はキメラは出来てない筈という判断でリジェクトされています。最初のネイチャーの査読者であった#1さんもキメラは出来てない筈と判断してリジェクトしているわけです。
できていると判断していながらリジェクトする商業雑誌は無いと思いますね。雑誌は売るのが目的ですから、こんな大発見を事実なのに掲載しないということはないでしょうね。事実でなくて掲載すると今度は信用失墜して売れなくなりますから、用心深く判断するわけです。
#1さんは要請を受けて、このキメラはESコンタミですよと意見しているわけです。だからこそ若山さんはそのときにESはなかったと書いておきなさいと指示しているわけです。
ところが今回は理研と笹井さんや丹羽さんのネームヴァリューが加わって編集部が信用したわけですよね。リヴァイズさせるという条件でコメントを書かせていますからこの#1さんもキメラは出来てないでしょうとは書けないんですね。ですからキメラ以外に関していろんな指摘をしている。
ここではまだででてきていませんがレターの査読では論文掲載をお勧めしないと書いています、二報同時だから再投稿できたのですから、レターは掲載しないということはやはり全否定なんですね。
キメラは出来てないでしようとはリヴァイズと要請している中では言えないので、別の解釈があるという指摘になる。それが死細胞の方をちゃんと数えなさいという指摘で、小保方さんはその指摘に応えてExtended Data Figure 1-g,hの実験を行ったのです。
こういうことを検討し始めると、直ぐに忘れがちですが、キメラが出来ていたら他の細かな雑多な証明は実は何も要らないのだという具合に証明法にもランクがあるということです。キメラが出来たらそれは多能性細胞です。Oct4遺伝子が発現したら多能性細胞であるかもしれない。なぜなら多能性細胞であるES細胞はOct4遺伝子を発現するからです。でもES細胞はキメラ証明されていますが、ES細胞でないものがOct4遺伝子を発現していたら、多能性細胞であるかもしれないから、次はどうするかというとキメラを実際に作ってみるわけです。何故ならOct4遺伝子を発現しているがまだキメラ確認されていないからです。
#1さんがどれほどおかしなコメントをしているかは理解できますよね。キメラは出来ていると認めた形でコメントしているのに言ってる内容はキメラは出来てないと言ってるのと同じことになるんですよね。
PI/Annexin-Vで死細胞を確認しなさいよと指示しているのですが、それを確認したら何が分かるのかというと、蛍光細胞が多能性細胞であるという可能性だけではないことが分かるでしょと言ってる。解釈はいろいろありうるのだよと言ってるんですが、ではあなたはなぜリヴァイズ要請コメントをしているのだというと、本心は別として、キメラが出来たということを認めたからでしようということになる。キメラが出来たことを認めると、この細胞は多能性細胞だということを認めたことになる。キメラは最終証明ですからね。
するとキメラが出来たことによって蛍光細胞の中の少なくとも何かが多能性細胞であったことが証明されたことになって、沢山あった可能性の一つ以外は否定されたということになるわけです。#1さんは自己矛盾を犯しているわけです。本来ならそれなら私は査読者を降りますというべきでしょうね。さもなければntESだということには気づかないまでもESのコンタミだから私は認めないと正直に書くべきなんですね。編集者との間に議論があったでしょうね。レフェリーとして残ったから苦しい書き方になっているわけです。桃子本では最後まで了承しなかったので編集部の責任で#1さんの意見は無視されてアクセプトされたといいますね。
GFP+というのは多能性細胞であるという判断です。無論キメラができているという前提です。
CD45+というのはまだ体性細胞である白血球だという判断です。
PI+は死細胞で核が赤く染色される。
g図はブルーがGFP+/CD45-/PI-ですから生きている多能性細胞という判断。オレンジはGFP-/CD45+/PI-ですから生きている白血球です。左図は酸浴未処理細胞群で全部生きている白血球です。d0からd7へ幾分生きている細胞数が減ってきているということです。縦軸は<% in total cells including PI+ cells>ですから死細胞も含めてFACS解析して生きている細胞だけを%表示したということです。右図は酸浴細胞群で、GFP-/CD45+/PI-とGFP+/CD45-/PI-の両方ができて来ている。酸浴直後のD0で既に20%の細胞が死滅していて、d7に向って生きている白血球が更に死滅していくが、生きている多能性細胞であるブルーの棒グラフが徐々に伸びて来るという図です。
この実験は#1さんがやらせた実験です。「施された細胞処置が遺伝子異常を引き起こす可能性に加えて、大量細胞死を引き起こしていることも明白である。著者らはその後の実験の流れに沿って、生き残った細胞に意識を向けて、図1に細胞数計測見積を示しているが、それは恐らく最善の実験ではない。著者らは細胞死の数量を実証し、かつ示し、そしてこれをOct4-GFP観測と同時にCD45陽性細胞を共染色することで実行しなければならない。これはPI/Annexin-Vを使ってできる簡単な実験である。」と言いましたよね。その結果、このブルーの伸びは「著者らは、体細胞系譜決定が遺伝子修飾に働きかけることによってのみ強力に変化させ得ると、又、その過程は自然には生じないと、読者を信じるように導いている。遺憾ながら私は同意しない。これは心筋細胞の部分的分化が心臓疾患治癒の原因となって以降のケースとは違う(Senyo et al.,2012 nature 事例を参照)。面白いことに、細胞系譜決定の動的平衡が癌幹細胞培養維持にも又示されてきている(Gupte et al.,2011 Cell 事例)。私見では、これらの事例は、著者らによって為されている主張に反して、哺乳動物体細胞は遺伝子修飾無しに、脱分化能力を持つことを示している。」ということを証明したでしょうか。
これは#1氏の論理飛躍のしすぎではないでしょうか。
でも、キメラは出来なかったという現在の知見からこれはどういう解釈になるのかというと、小保方さんの酸浴細胞は大半死に絶えながらも生き残った細胞は丹羽さんの検証実験によれば内在性Oct4はほとんど発現しないのに、マーカーであるGFPだけが強く発現する細胞が残ってくるということが分かったわけです。と同時に、亜致死下の細胞がすべて微量の内在性Oct4を発現していることも小保方さんによって証明されていましたね。

a, Immunostaining for pluripotent cell markers (red) in day 7 Oct4-GFP+ (green) clusters. DAPI, white. Scale bar, 50 μm.
a,7日目のOct4-GFP陽性(緑)クラスター内の多能性細胞マーカー(赤)の免疫染色。 DAPI、白。スケールバーは50μm。
左上です。これは免染結果ですから定性分析です。すべての細胞が微量に内在性のOct4蛋白質を発現しているんです。丹羽さんの漏れ出しの表は定量分析ですからGFP遺伝子の発現量に対しては内在性Oct4遺伝子はゼロになっている。

(a)Q-PCR analysis of the low-pH treated liver cells cultured for 7 days. Liver cells were prepared from 7-day old GOF mice and treated with either ATP or HCl, or without stressor. RNA samples were prepared from all cells in the wells at day 7 of culture and the relative expression levels of Gfp (derived from GOF Tg) and Oct3/4 (derived from the endogenous Pou5f1 allele) to Gapdh were indicated with standard deviation. The expression levels in control ES cells carrying CAG-GFP Tg were set at 1.0.
(a)7日間培養した低pH処理肝細胞のQ-PCR分析。肝細胞を7日齢のGOFマウスから調製し、ATPまたはHClのいずれか、またはストレッサーなしで処理した。培養7日目にウェル中の全細胞からRNA試料を調製し、Gfp(GOF Tg由来)およびOct3 / 4(内因性Pou5f1対立遺伝子由来)のGapdh<訳注:glyceraldehyde-3-phosphate dehydrogenase;GAPDH mRNAはさまざまな生理状態において発現量が変わらないと考えられている>に対する相対発現レベルを標準偏差で示した。 CAG-GFP Tgを保有するコントロールES細胞における発現レベルを1.0に設定した。
Oct4-GFPを仕込んだマウス由来のES細胞を使っていませんね。CAGマウスのES細胞です。その発現量をGapdhの発現量と比較してその比を1としている。対して酸浴細胞はOct4-GFPですからCAG-GFPとOct4-GFPの発現量は発現する時には同じ量発現するという前提になっているんですが、そのようなことは分かってないのではないですかね。一番いいのはOct4-GFPを持つマウスのES細胞をコントロールとして使うのがいいですよね。
しかし、焦点の問題として、Oct4-GFPが発現しているのに内在性Oct4遺伝子は発現してないということがあるのだという事実は証明されているわけです。ただ、その量はまずCAG-GFPの発現量が基準になっているのと、もう一つは相沢さんの指摘している酸浴細胞のGapdh値が動いているというところに信頼性が欠如しているわけですね。
ですから、小保方さんの酸浴細胞が何であるのかということはまだまだ解明されてはいないのです。でも、相沢さんが小保方さんと雑談中に聞き出したこれまでの経緯の話によって、キメラは若山さんの訳ありの方便の嘘だと分かってしまったので、急いですべてを有耶無耶にして終わらせたというだけの話です。論文通りのキメラではないのだと分かったのです。
次に、h図はまず赤色のPI+/Annexin+は完全死細胞で、オレンジ色のPI-/Annexin+は初期アポトーシス、ピンク色のPI-/Annexin-は正常細胞です。<GFP-/CD45+>は白血球細胞集団だと思いますが、<GFP+/CD45- Total>の意味がよく分からない。何と何のトータルなのか。特に左図の未処理細胞には<GFP+/CD45->はないことにg図でなっていますから、トータルしても同じはずですが、<GFP-/CD45+>自体より余計に細胞死している。ここは良く理解できません。
本文の続きです。
>
>著者らは観測されている過程が実際に細胞転換に思えるのか、腫瘍化なのかを定義するように努力しないといけない。比較はOct4-GFP陽性細胞と非処理CD45陽性細胞との間で行われているが、最善の内部コントロールとしては処理中にできてくるOct4-GFP陰性細胞の集団である。
非処理のCD45+細胞(白血球細胞集団)から酸浴処理後にOct4-GFP陽性細胞ができると同時に生きたままのOct4-GFP陰性細胞も残っているが、それはCD45陽性細胞(白血球細胞集団)ではないのかという疑義ですね。
本文は続きます。
>
>図1bは適正に示されていない。すべてのコントロール細胞を含む適切な識別手法が示されなければならない。同様に、それぞれのパネルはきちんとラベルしておかないといけない。読者はなぜ非処理のCD45陽性細胞が(三日目の)CD45の減少を示すことになったり、7日目までの完全に非処理の独立した細胞集団ですら減少することになるのか戸迷うことになる。又、著者らは、過剰造血細胞系列と前駆細胞を含むCD45陽性細胞としての出発点の細胞集団の特性定義を求められている。
どうやらオリジナルの図1bは最終版の図1dのようですね。
d, GFP+ (green) and GFP− (yellow) cell populations (average cell numbers per visual field; ×10 objective lens). n = 25; error bars show average ± s.d.
d,GFP +(緑)、GFP-(黄色)の細胞集団(視野当たりの平均細胞数、×10対物レンズ)。 試行数= 25;エラーバーは、平均±のs.d.を表示。
D0というのは酸浴直後ですから、ここには酸浴前のデータがないんですね。そもそも酸浴前のCD45+細胞の定義を求めているわけで、その後に今度は酸浴以降に常に幾分生き残っている、ここではOct4-GFP-とされている細胞集団の定義を求めている。つまりそれらはGFP発現がないのはいいとして、そもそもまだCD45抗原を持って維持しているのかということですね。最終図は以下です。

オリジナルでは以下のaがあって後に今のdがbとして提示されていたのでしょうかね。最終形ではその間に今のb,cが挿入された。

☟

☟

☟
この部分に関する本文は以下ですね。
Low pH triggers fate conversion in somatic cells
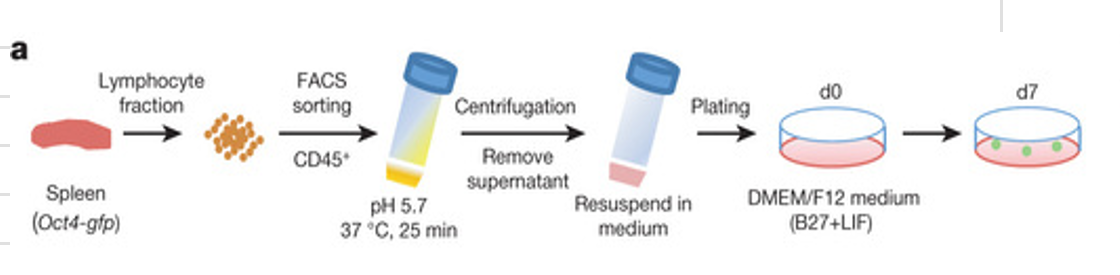
CD45+ cells were sorted by fluorescence-activated cell sorting (FACS) from the lymphocyte fraction of postnatal spleens (1-week old) of C57BL/6 mice carrying an Oct4-gfp transgene, and were exposed to various types of strong, transient, physical and chemical stimuli (described below). We examined these cells for activation of the Oct4 promoter after culture for several days in suspension using DMEM/F12 medium supplemented with leukaemia inhibitory factor (LIF) and B27 (hereafter called LIF+B27 medium).
Among the various perturbations, we were particularly interested in low-pH perturbations for two reasons. First, as shown below, low-pH treatment turned out to be most effective for the induction of Oct4. Second, classical experimental embryology has shown that a transient low-pH treatment under ‘sublethal’ conditions can alter the differentiation status of tissues. Spontaneous neural conversion from salamander animal caps by soaking the tissues in citrate-based acidic medium below pH?6.0 has been demonstrated previously.
Without exposure to the stimuli, none of the cells sorted with CD45 expressed Oct4-GFP regardless of the culture period in LIF+B27 medium. In contrast, a 30-min treatment with low-pH medium (25-min incubation followed by 5-min centrifugation; Fig. 1a; the most effective range was pH 5.4–5.8; Extended Data Fig. 1a) caused the emergence of substantial numbers of spherical clusters that expressed Oct4-GFP in day-7 culture (Fig. 1b). Substantial numbers of GFP+ cells appeared in all cases performed with neonatal splenic cells (n = 30 experiments).
The emergence of Oct4-GFP+ cells at the expense of CD45+ cells was also observed by flow cytometry (Fig. 1c, top, and Extended Data Fig. 1b, c). We next fractionated CD45+ cells into populations positive and negative for CD90 (T cells), CD19 (B cells) and CD34 (haematopoietic progenitors), and subjected them to low-pH treatment. Cells of these fractions, including T and B cells, generated Oct4-GFP+ cells at an efficacy comparable to unfractionated CD45+ cells (25–50% of surviving cells on day 7), except for CD34+ haematopoietic progenitors, which rarely produced Oct4-GFP+ cells (<2%; Extended Data Fig. 1d).
低ペーハー刺激による体細胞の運命転換
CD45+細胞は蛍光活性化細胞選別器(FACS)によって、Oct4-GFP導入遺伝子を保有するC57BL/6マウス(1週齢)の出生後の脾臓のリンパ球画分から選別された。そして様々なタイプの強い一時的な、物理的、化学的刺激に曝された(下記参照)。我々は白血病抑制因子(LIF)及びB27(以下LIF+ B27培地と呼ぶ)を補充したDMEM/ F12培地を使用した懸濁液中で数日間培養後Oct4プロモーターの活性化についてこれらの細胞を調べた。
さまざまな撹乱の中で、我々は2つの理由から低pHによる撹乱に特に興味を持った。以下に示すように、まず、低pH処理は、Oct4の誘導のために最も効果的であることが判明した。次に、古典的実験発生学は「亜致死」条件下での一過性低pH処理が組織の分化状態を変化させることができることを示している。 pH6.0以下のクエン酸ベース酸性媒体中に組織を浸すことによってサンショウウオのアニマルキャップからの自発的神経変換が以前に実証されている。
刺激に曝さないままだと、CD45でソートされた細胞はどれもLIF+ B27培地での培養期間にも関わらず、Oct4-GFPを発現しなかった。対照的に、低pH培地での30分間処理は(25分間の保温期間に5分間遠心分離<図1a>、最も効果的な範囲はpH5.4から5.8であった<拡張データ図1a>。)7日の培養においてOct4-GFPを発現する多くの数のスフィア状クラスターの出現を引き起こした(図1b)。 GFP 陽性細胞の大量の数は新生児の脾臓細胞で実行されたすべてのケースで現れた(30回の実験)。
CD45陽性細胞を使ったOct4-GFP陽性細胞の出現はまたフローサイトメトリー装置によっても観察された(図1c、最上部、及び拡張データ図1b、c)。我々は次にCD45陽性細胞をCD90(T細胞)、CD19(B細胞)およびCD34(造血前駆細胞)の陽性と陰性の集団に分画し、それらを低pH処理に供した。T細胞とB細胞を含むこれらの分画細胞は、Oct4-GFP陽性細胞をめったに作らなかったCD34陽性造血前駆細胞を除いて(2%以下;拡張データ図1d)、未分画CD45陽性細胞(7日目に生存している細胞は25〜50%)に匹敵する効果でOct4-GFP陽性細胞を発生した。
最後のところに脾臓から取得した白血球特異的分化抗原であるCD45を発現している細胞集団を更に分画して、CD90(T細胞)、CD19(B細胞)およびCD34(造血前駆細胞)とそれ以外のつまりCD45細胞の4つに分類したが、造血前駆細胞はGFPを発現する細胞にならなかったと書いている。
これは実際に小保方さんが#1さんに言われて行った実験ですが、これは大きな確認で造血前駆細胞は幹細胞からT細胞やB細胞に分化する直前の状態ですから幾分まだ多能性を持っている可能性がある。それはOct4-GFPを発現しないと確認しているわけです。もっとも前駆細胞をキメラ胚に入れてもキメラは出来てきません。これらの体性幹細胞はすでに全能性は失ってるんですね。ですからこの結果は別に小保方さんが強弁のために嘘をついているわけでもないんですね。単なる事実なんですね。
でも、結果的には小保方さんの細胞は論文通りのキメラなんてできてませんから、大本の嘘に騙されているわけです。でもそれは嘘だったのだとすると小保方さんのこの確認は何を意味しているかというと、GOFマウスに仕込まれているOct4-GFP人工遺伝子構造は他の細胞では丹羽さんの発見したGFPの遺伝子異常発現を引き起こす設計になってしまっているが、前駆細胞ではその異常は起こらないのだという類の一つの事実の発見になるんですね。するとOhoboさんたちが何故細胞を虐めると自分たちの設計した人工遺伝子が正常に発現しなくなるのかを突き止めるための重要なデータになりうるんですね。基礎研究というのはそういうもので、知らなかった事実をとにかくたくさん集めることで、それらが何かの時に新たな大きな発見に繋がることが千三つあるんですね。だからこういう事件になりましたが失敗なんて研究の常態ですから、本来は世間がとやかく言うようなことではないのです。
(2022/6/16)
酸浴細胞でなぜ内在性のOct4遺伝子は発現しないのにOct4-GFP人工遺伝子が発現するのかという問題は丹羽さんによってポンと投げ捨てられたのち関係者には放置されたままになっている。その理由は、これが事件の収拾策の中にあるからなんですね。事件が無かったとしたら、どうしてそんなことが起きるのだろうと興味深く研究者は思って、いろんな研究をした結果、論文が出るのが当たり前ですね。でも出てません。事件になってますから出たら世間が騒ぐから分かる。だから実際、出てないんですね。また、事件になってなければ論文は出たでしょうが、世間は気づきもしなかったでしょう。
要するにこの分野の研究界のタブーになってしまっているわけです。事件を蒸し返さないためですね。他にやることはいくらでもありますからね。それは誰にでも理解可能ではないか。
遺伝子というのは開始コドンがあって終始コドンで読み取りが終わります。で、読み取りの開始はどういう仕組みで起きるのかというと、構造遺伝子の前方にプロモーター領域という配列があってその部分に他の要因で作られた蛋白質が結合することが契機になって、いろんな種類があるが、代表的なものとしては、それまで読み取りができないようになっていた部分が取り外されることで読み取りが開始される。
GOFマウスには普通の遺伝子だけでなくGFP人工遺伝子が組み込まれている。
①[Oct4遺伝子プロモーター領域配列]+[Oct4構造遺伝子]---(内在性遺伝子)
②[Oct4遺伝子プロモーター領域配列]+[GFP構造遺伝子]---(人工遺伝子)
常識的に考えると[Oct4遺伝子プロモーター領域配列]に働きかける蛋白質が核内に入ると当然①も②も同時に発現しますよね。②の蛍光発現は①の発現を保証している。だからこそその便利さからGOFマウスとして販売されているんです。この蛍光遺伝子発現が無かったら免染で確認しないといけないですがマウスを殺さないと免染は出来ません。②があると生きているままで確認できるわけです。
ところが小保方さんが細胞を酸浴させるという乱暴な処置をしたら①は発現してないのに②だけ発現する現象があるという事実を丹羽さんが発見した。理屈上そんなことはあり得ませんよね。ならば理屈が間違っているんですね。誰が考えてもこんな単純な論理に間違いはありませんから、間違いは前提にあるに決まっている。
ど素人の推測では②の概念図に間違いがあるのだと思われますね。DNA配列はほとんどの部分がジャンク配列で無意味な配列が間にたくさんある。でも、それがどういう働きをしているのかはまだまだ解明途上なんですよね。分かってないんです。②は恐らく以下なんです。
②[Oct4遺伝子プロモーター領域配列]+<イントロン>+[GFP構造遺伝子]---(人工遺伝子)
①は例えば恐らく以下なんです。
①[Oct4遺伝子プロモーター領域配列]+<イントロン>+<イントロン>+[Oct4構造遺伝子]---(内在性遺伝子)
つまり人工遺伝子を作る時に切り取ってきた部分だけの働きと自然の配列には当然違いがあって、現在の知見ではそのイントロン部分には働きがないとされていて、実際に普通の状態では働かないが酸浴という特殊環境でイントロン部分に何らかの働きが生じて②だけが発現するというような可能性ですね。
すると、小保方さんが査読要請に従って前駆細胞ではGFP蛍光しないという事実を発見していることが、何か前駆細胞特異的な他の蛋白質の介在が漏れ出しを防いでいるのだと推定すると、①と②とのイントロン部分の違いの何がそうさせているのかの原因解明に寄与しうるデータとなるわけです。
ですから、常識で考えて、実験事実は事実として正確に報告しなければならないんですね。お金かかっていますから、事実を知るのに只ではないんですから、全人類が働いたお金の一部を研究に回して、ニュートン曰くところの浜辺の全砂粒の中の一つの砂粒程度に関して知るのに人類は何百年を使っているわけです。事実の集積があって初めて法則の構築が可能になる。具体的に何も分かってないのに法則が頭脳に浮かぶということはあり得ないので赤ん坊は何も知らないんですよね。
基礎研究って大変ですね。もっとも世間全員誰でも生きて行くのにはそれぞれ大変な思いをしてますけどね。
本文です。
>
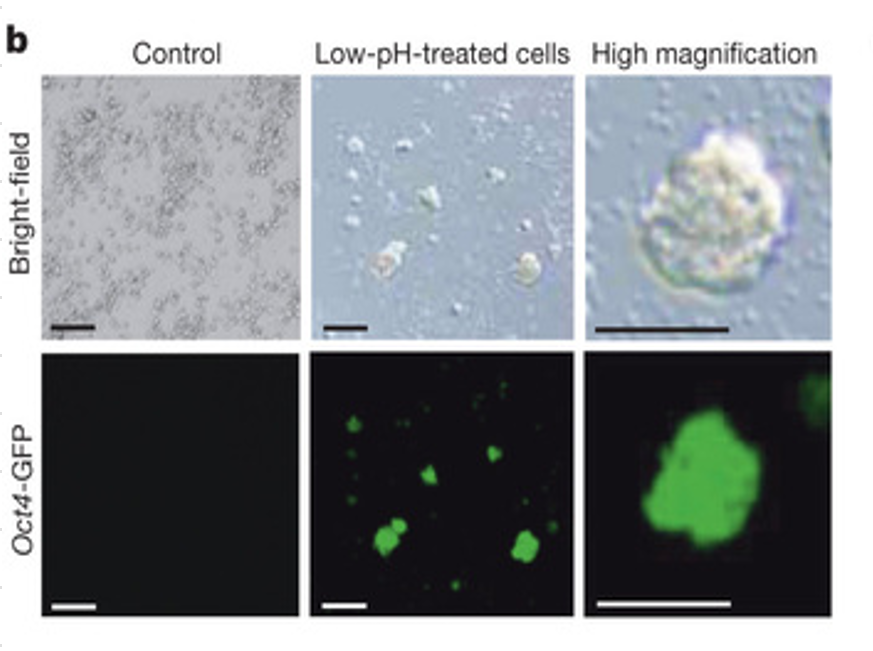
>著者らはSTAP細胞が小さいということによって何を主張したいのか? 何の関係があるのか? 何故? 葉状仮足って何? 著者らは、これらの実験の背後にある疑問や仮説が何に関するものなのか、又何が結論として引き出されてくるのかを解説すべきだ。査読者である私にはこれらの観察結果の関連が示されているとは見えない。代わりに、クロマチン沈降実験と脱メチル化の情報提供をすると確かにより興味深いのではないか。必ずしもゲノムワイドプロファイリングまでは必要ではないが、簡単なヒストン修飾確認、かつ又は、脱メチル化確認実験があるととても論文の質が高まるのでないか。
ちょっと私の英語力では誤訳しているかもしれませんね。最初の方はいいんですね。細胞が小さいというのは事実を指摘しているだけです。当初は酸浴しているので中の水分が出て痩せているのではないかとも思っていたかもしれませんね。手記には細胞膜の損傷が関係してないかとも考えていたりしていたようですね。私は単にリンパ球の大きさが他の細胞種とは違ってかなり小さい細胞だったからだとみています。後に、若山さんの嘘として私は大きさの違いは分かったはずだとして以下の図を示しましたが、この査読段階では若山さんが胚盤胞にインジェクトする時に大きさの違いは分かったはずだなどと小保方さんが考えるわけもありませんから、後の事件化後の推測とは無関係で只小さいという事実を書いているだけです。


小保方さんはオリジナルの原稿にg図をつけていたのでしょうね。

査読者はこういうのを見て説明が足りないとクレームしているのでしょうね。
葉状仮足に関しては最終版の動画のレジェンドに以下のようにあるんですね。これは今でも見れます。本文は有料になっていますがサプリはダウンロードできます。私も以前これを見て笹井さんはここまでは見てやっていないのだろうなと思いましたね。リンパ球はアメーバではありませんから仮足運動はしませんので、笹井さんが見てやっていたら、これは焦点が合わせられていないから見えないが、マクロファージが死細胞を食べて動いているから引きずられているのだということはすぐに分かったはずです。小保方さんはリンパ球そのものが動いていると感じていたわけですが、深くも考えずに見たままに書いている。見たままの描写としては間違ってはいないわけです。
>>
査読者にいろいろと言われてこの部分に関する最終稿の本文は以下になった。
>>
Low-pH-treated CD45+ cells, but not untreated cells, gradually turned on GFP signals over the first few days (Fig. 1e, Supplementary Videos 1 and 2 and Extended Data Fig. 2a), whereas CD45 immunoreactivity became gradually reduced in the cells that demonstrated Oct4-GFP expression (Fig. 1f and Extended Data Fig. 2b). By day 5, the Oct4-GFP+ cells attached together and formed clusters by accretion. These GFP+ clusters (but not GFP− cells) were quite mobile and often showed cell processes on moving (Supplementary Video 1).
The Oct4-GFP+ cells demonstrated a characteristic small cell size with little cytoplasm and also showed a distinct fine structure of the nucleus compared with that of parental CD45+ lymphocytes (Fig. 1g). The Oct4-GFP+ cells on day 7 were smaller than non-treated CD45+ cells (Fig. 1g, h and Extended Data Fig. 2c) and embryonic stem (ES) cells (Fig. 1h), both of which are generally considered to be small in size. The diameter of low-pH-treated CD45+ cells became reduced during the first 2 days, even before they started Oct4-GFP expression (Fig. 1f), whereas the onset of GFP expression was not accompanied by cell divisions. Consistent with this, no substantial 5-ethynyl-2′-deoxyuridine (EdU) uptake was observed in the Oct4-GFP+ cells after the stressor (Extended Data Fig. 2d).
CD45の免疫反応性がOct4-GFP発現の証明された細胞の中で徐々に減少したのに対して(図1f及び拡張データ図2b)、未処理細胞で無い低pH処理されたCD45陽性細胞は最初の数日間で徐々にGFPシグナルを発し始めた(図1e、補足動画1、2と拡張データ図2a)。5日目では、Oct4-GFP陽性細胞が互いに密着し、集積によってクラスタを形成した。これらのGFP陽性クラスター(ただし、GFP陰性細胞ではない)は大変可動で、しばしば動いている細胞過程を示した(補足ビデオ1)。
Oct4-GFP陽性細胞は細胞質のほとんどない特徴的な小細胞サイズを示し、また親のCD45陽性リンパ球のそれと比較して異なる核の微細構造を示した(図1g)。7日目のOct4-GFP陽性細胞は非処理CD45陽性細胞と胚性幹(ES)細胞(図1h)よりも小さかった(図1g、hおよび拡張データ図2c)。どちらも一般的にサイズの小さい細胞であると考えられている。低pH処理したCD45陽性細胞の直径は、GFP発現の発生が細胞分裂を伴わなかったにも関わらず、それらがOct4-GFP発現を開始する前であってすら、最初の2日の間で小さくなりだした(図1f)。これと同調して、いかほどの5エチニル-2'-デオキシウリジン(EdU)の取り込みもそのストレス因子後のOct4-GFP陽性細胞の中に認められなかった(拡張データ図2d)。
サイズに関してはCD45+細胞とES細胞のどちらも一般的には小さい細胞だと書いていて、酸浴後のSTAP細胞は細胞質が減ってもっと小さいと強調していますね。でも私が指摘しているように、小保方さんはサイズバーを正確に合わせていません。これは三つを並列して見やすくするためで、他方でキメラができていますから安心しているのでしょうが、上述しているように実験は事実通りに報告しないといけません。サイズバーを合わせると私が作った図になる。
重大なのは実寸比較していたら、単にES細胞がCD45+細胞より大きいということだけでなく、STAP幹細胞がESの大きさになっていることに気づけたはずなんですね。若山さんがntES化しているからES の大きさになっている。
小保方さんは<The Oct4-GFP+ cells demonstrated a characteristic small cell size with little cytoplasm and also showed a distinct fine structure of the nucleus compared with that of parental CD45+ lymphocytes (Fig. 1g). The Oct4-GFP+ cells on day 7 were smaller than non-treated CD45+ cells (Fig. 1g, h and Extended Data Fig. 2c) and embryonic stem (ES) cells (Fig. 1h), both of which are generally considered to be small in size.>と書いてますよね。こうであろうという自分の仮説の言葉に引きずられているから事実が事実通りに見えてないんです。言葉の力というのは大きいんですね。一般にも「思い込み」といいますね。視覚という直覚は言語と強く結びついていて、見えているものを関連付けて理解しようと努力するんですが、その時に自覚がないと「思い込んだ」理解のために不都合なものが見えなくなるんですね。
私が誤訳かもしれないと思うのは以下の部分です。
>>
代わりに、クロマチン沈降実験と脱メチル化の情報提供をすると確かにより興味深いのではないか。必ずしもゲノムワイドプロファイリングである必要まではないが、簡単なヒストン修飾確認、かつ又は、脱メチル化確認実験があるととても論文の質が高まるのでないか。
以下のInstead以下です。前の疑問とどういう繋がりになっているのかよく分からない。

Insteadの意味が前述した疑問に関して、そんな比較をするよりもChIPシーケンスや脱メチル化確認をした方がいいという意味なら、これはちょっと変なんですね。オリジナル投稿論文にはメチル化実験が無かったということになるんですよね。でも、この実験は最初のネイチャー投稿時に添付されていたデータだと桂報告書に書かれていましたよね。スライドです。

本文です。
私の査読書読み取りでは、サイエンスベースの12/25ヴァージョン原稿を笹井さんが書き直してやった時のオリジナル投稿原稿には、最初のネイチャー、セルにはあったこのメチル化実験結果は無かったという解釈になる。何か誤解誤訳しているかなあ。
桂報告書の書き方でも「この図は 2012 年 4 月の Nature 投稿原稿、Cell 投稿原稿でも使われ、最終的に Article Fig.2c として発表された。」としていてサイエンス原稿に関して触れてませんね。小保方さんは「これでは論文に使えないね」と言われてモチャモチャやったことを認めています。それは最初のネイチャー論文に掲載されて、セルまでは使われたが、小保方さんはサイエンスではもういいだろうと思って外していますよね。この時点で不正は無くなっていた。そしてサイエンスもリジェクトされたのでサイエンスベースでの改訂版を作製していて、それを理研に呼び戻された時に日本への飛行機の中で12/25と日付を打ったんですね。そしてそれを笹井さんが書き直すことになった。書き直しのときに論文原稿に存在してないものは二人で討論はできませんね。するとこのとき無かったものを付けさせる契機になったのは査読者のこのコメントだということになるのですが、この#1さんは最初のネイチャー論文の査読者ですから、そこにメチル化実験結果が添付されていたのを知っているんですよね。
他方で「STAP 幹細胞についてもメチル化解析が行われ、この結果は 2013 年 3 月に投 稿された Letter 原稿 Fig.3 として初めて提示され、最終的には Article Extended Data Fig.8d として発表されている。」と書かれていることによって、レター論文に関して笹井さんが書いた3月投稿時に既にメチル化結果が添付されていたとしているのが本当なら、アーティクル側のメチル化実験結果もつけられていたと考えるのが普通です。ただし、誤解のないようにしなければいけないのはSTAP幹細胞はntESですから脱メチル化しています。仮にES細胞でも脱メチル化している。捏造の必要はありません。対してSTAP細胞は小保方さんが自分で作った段階で行っていますから、脱メチル化してないんですね。でもキメラは出来たと思い込まされているのでモチャモチャやったんです。ESでchimera捏造なんてしてたらモチャモチャすることも無いと分かりますよね。
さあ、査読者はアーティクル論文に脱メチル化結果があったらいいのにと言ってるのなら、最初はついてなかったことになる。私の誤読なんですかね。
そもそもこれをつけた方がいいのは若山さんの都合だと考えるとこの査読者と通じていないかと私が最初に疑った理由も理解できると思います。
そしてでも、レターにはついていたようなんですよね。
小保方さんは酸浴細胞はモチャモチャやったので外したが、レターは事実通りだったから添付したのでしょうかね。
特許書類ではACCsになっていて、これはネイチャー、セルの呼び名です。サイエンスではSACsですね。

対してリトラクトされたレター論文にあった図はヴァカンティ氏は特許書類から現在全部外しているのですが、そこに「STAP 幹細胞についてもメチル化解析が行われ、この結果は 2013 年 3 月に投 稿された Letter 原稿 Fig.3 として初めて提示され、最終的には Article Extended Data Fig.8d として発表されている。」とされた同じ図がアーカイヴとして残されている。無論本文では抹消されている。

STAP-S CELLSとされているから削除されたんです。リトラクトされたネイチャー論文にあるものだから削除した。つまり三誌には無かったということで、これは三誌はキメラが出来たという論文で、幹細胞化論文は若山さんの研究ですからもともと無いんですね。
すると、このデータは若山さんが小保方さんに渡した幹細胞化のデータの中に含まれていたということになる。サイエンスベースの12/25ヴァージョン原稿の中にはもうメチル化実験の図は外されているが、若山さんの研究データの中には原稿はまだできてないがデータとして存在していたわけです。笹井さんはレター原稿を書いたときにそれを図として取り込んだ。しかし、アーティクル側に同じ分析がないことには気づかなかった。とすると、#1さんはレター原稿との比較はしなかったのかということになる。
無論若山さんのデータの中にあったとはいえ、実際に確認実験を行わされているのは小保方さんでしょうね。若山さんは酸浴細胞の脱メチル化実験を小保方さんにさせたと同時に、STAP幹細胞の脱メチル化実験も同時にやらせているんですよね。自分の酸浴細胞は脱メチル化してないのに、自分にはできないが若山さんの培養したSTAP幹細胞は脱メチル化していて、かつどちらもキメラが出来たと言われているわけです。その両方の結果を見て小保方さんは何故だろうと思わなかったのでしょうかね。
こういう問題の出し方というのはとてもテスト臭くないですかね。








| オリジナル・メッセージ |
| 件名 : ネイチャー誌■■■に関する決定 |
| 送信者 : ■■■ |
| 送信日時 : 2013年04月04日(木) 22:44:04 |
| 宛先 : ■■■ |
| 2013年4月4日 |
| >*共著者達に転送したい場合はこのEメイルのあなたの著者ホームページへのリンクを消したことをご自身で確認してください。 |
| > |
| >拝啓 小保方博士殿 |
| > |
| >御健勝のことと存じます。"体細胞の多能性への刺激惹起性運命転換"と題されたあなたの論文は現在3人のレフェリーによる査読を受けており、そのコメントは以下に添付されています。我々もそうですが、彼らもあなたの大きな可能性を秘めた業績に関心を抱いている半面、ネイチャー誌に掲載する判断の前に、我々の視点では御回答願わなければならない重要な問題点を指摘しています。 |
| > |
| >これらの批判に答える更なる実験データを載せた改訂版を戴けると幸甚です。(何か似た論文がネイチャー誌や他の場所に同時期に書かれている場合は別です。) |
| > |
| >改定論文は以下のURL(www.nature.com/nature/authors)に置かれている我が誌の書式指示及び出版規則に従ってください。改定後の論文は以下のリンクを使って提出してください。 |
| > |
| >■■■ |
| > |
| > |
| >*このURLはあなたの個人名ホームページと論文に関する関連情報にリンクしており、あなたが提出に使ったり、我々のために確認したりすることができます。もし共著者達にこのEメイルを転送したい場合はまず最初にあなたのホームページへのリンクを消してください。 |
| > |
| >同時に我々はあなたに我々のレフェリーコメントをよくお読みいただけることを望んでいます。相談したいことがあったらご遠慮なく私の方に連絡ください。 |
| > |
| >敬具 |
| > |
| >■■■ |
| > |
| >■■■ |
| >■■■ |
| >ネイチャー |
まず題名はオリジナルでは<Stimulaus-Triggered Fate Conversion of Somatic Cells into Pluripotency>であった。最終稿のタイトルも同じですが大小文字の使い分けを変えていますね。<Stimulus-triggered fate conversion of somatic cells into pluripotency>となっている。何故かは分かりません。
レフェリーのコメントはそもそも専門的なことなのでど素人には分かりにくいのですが、何よりもオリジナルの論文の文面と図が無いので何を言わんとしているのかの推測がとても難しい。専門であっても結構むつかしいのではないかと思いますが、門外ですから常識の範囲にないことの推測は大変です。
パラグラフごとに翻訳していきましょう。
>レフェリー・コメント
>
>リフェリー #1 (著者への所見)
>
>この現在の論文では、著者らは、多能性へ向けた細胞核のリプログラム過程での細胞への刺激の役割を重要視している。これはとても興味深く、恐らく革新的な論文である。しかし、結論に導く説明とデータは幾分推論に過ぎず、かつ、一部は手始めの段階である。
>
ちなみにこの#1は2011年に三誌論文の最初に投稿したネイチャーの査読者と同じ人だと桃子が言ってますね。桃子本308P。
>>
まず目を引いたのは、ネイチャー編集者のコメントの変化だった。
初稿当時には「論文は掲載できない」と淡々とした文面で伝えたのに対し、再投稿時には大きな関心を示し、六カ月以内に追加実験をしたうえで、改訂するよう強く勧めて、「改訂版の原稿をぜひ見たい」と記した。これを読んだある大学教授は「熱狂的といってもいいような反応、初稿当時とあまりに違う」と驚いた。
一方、査読者たちは、「発生生物学や幹細胞生物学の転換を迫る発見」「特筆すべき発見」などと賞賛しつつも、問題点や疑問点を多数挙げていた。
人数は一人増えて三人になっていたが、一人目の査読者が初投稿時の一人目と同じ人物であることは明らかだった。アーティクル論文に関するコメントの最初の一文が全く同じな上に、同様のフレーズが何度も登場したからだ。初投稿時は前向きな表現が見当たらなかったが、再投稿時は「非常に興味深く、画期的な可能性を秘めている」と評価する言葉があり、「結論を支えるデータと説明は非常に推論的で予備的だ」という一文は「結論を支えるデータと説明は少々推論的で、いくつかのケースでは予備的だ」と弱められていた。
おそらく同じメンバーがレター論文も査読しており、一人目の査読者はレターに関するコメントの最後にこう指摘した。-----著者達はSTAP細胞が次世代の再生医療の材料として有望だと主張するが、両論文中にはES細胞やiPS細胞と比較してSTAP細胞の(材料としての)品質を評価する実験が一つもない。ゲノムレベルでの詳細な解析をしない限り、この論文の掲載を勧められない。私は現在のデータを疑いはしないが、プロセスを要約すれば「魔法のような(magical)」アプローチだ-----。
編集からのメッセージ中で中央に大きく墨消しのある部分に桃子の書いている<六カ月以内に追加実験をしたうえで、改訂するよう強く勧めて、「改訂版の原稿をぜひ見たい」と記した。>内容があったのでしょうね。なぜこの部分が墨で消されているのかも流出元の配慮を推測する上での疑義の一つです。桃子が見た時にはこの文面は消されていないのですから、
①桃子が消したか、
②私がそのホームページからの入手した慶応大学の吉村氏が消したか、
③吉村氏がこんな流出してはいけない筈の査読書がネットに流されていると紹介したもとの書類に墨消しが既にあったのか、
のいずれかです。桃子は本に書いてますから①はない。②は意図的に吉村氏が流したのなら、ネイチャー誌の積極姿勢は隠してあげようと配慮したことになる。③の場合は、桃子は拡散してはいけない多くの入手書類を意見を聞くという名目で何人かの学者に配布している。その中の誰かが意図的に流したのならこの部分を隠す理由は弱いんですね。言うまでもなく吉村氏は遠藤論文の謝辞の中に出て来る仲間ですから、どうも②の香りがしますかね。
この第一レフェリーが誰であるかに関しては全世界的にもとても狭い社会なので仲間内では推測可能なのではないでしょうかね。三誌論文がすべてリジェクトされた時に若山さんは以下のように言ったとされている。手記103P。
>>
春から投稿されていたスフェアの論文は、残る最後の三大誌であるサイエンスからも、レビューワーにはまわるものの2011年8月21日には不採択の連絡が届けられた。若山先生から、「一度不採択になった論文は通常もう一度投稿することができないが、幹細胞株化の論文と2報同時に投稿すれば、再考されると思う。三大誌3つのレビューワーコメントの中で、ネイチャーからのコメントが一番やさしいので、ネイチャーに現在投稿を試みている論文と幹細胞株化の論文を2報同時投稿しよう」と提案され、幹細胞株化の論文の執筆とともに現在投稿中の論文に対するネイチャーからのレビューワーコメントへの反論を用意するように指示された。また、ES細胞の混入による可能性を否定するために、「ES細胞は同時に培養していなかった」という記事を原稿に加えることも指示された。
最初のネイチャー誌投稿は2012年の4月ですから投稿論文の実験中に培養されているES細胞は小保方さんの持っている学生のES細胞以外は無いのですが、4月末から5月初旬にFLSのジャームライントランスミッション結果が出ていて、サイエンスには二報同時論文と同じ結果図が載せられている筈ですからそこにコントロールESキメラのジャームライントランスミッション結果も載せられていたはずです。小保方さんはキメラを作れませんから以下のデータのESキメラも若山さんが実験している筈のものです。
サイエンスは8月21日にリジェクトされたことは分かっていますが何時投稿したのかを理研は発表していません。全部分かっている筈ですね。桃子は全部の書類を入手している。20日前だとしてみましょう。
8月1日だとしてみるのです。ESのデータは当然その時には出ているわけです。まず生きて生まれた数が書かれていて、成長できた数も書かれている。adulthoodというのは生後何日までを見るのかは知りませんので、生殖能力を得るのは生後50日後とされていますが、ここでは10日は生きて生育していたという意味にとっておきましょう。この場合生まれたのは7月20日前後ということになる。この前に生まれて来るのに20日掛かります。交配は7月1日前後です。交配できるようになるのに親のキメラマウスは50日掛かります。逆残すると親のESキメラマウスが生まれたのは5月10日前後になる。この時点で既に若山さんの事後MTA の細胞リストに記載されたコントロールES細胞の樹立日が5月25日になっているのは嘘だということになってしまうんですね。これはadulthood期間の10日仮定をゼロ日にしても間に合っていませんね。更に、キメラを作るのにインジェクト後20日かかる。インジェクションは4月20日前後だということになるのです。
若山さんの事後MTAの細胞リストは2014年4月1日付で提出されている。ここにはコントロールES細胞の樹立は2012年5月25日とされている。しかし、その後、桂報告書が2014/12/25に公開された時、その細胞リストには2012/4/19の樹立開始と書かれていたのです。本文中には以下のようにある。
>>
4)他の細胞株における遺伝的不均一性 この遺伝的不均一性は、129 CAG-GFP マウスに由来する他の細胞株にも反映していた。 若山氏により 129B6F1CAG-GFP マウスの独立した胚より複数の受精卵 ES 細胞株が樹立 されているが(129B6 F1ES1~6、2012 年 5 月作製)、いずれも第 6 染色体中程に B6 ホモ 領域を有していた。
細胞リストの樹立日は培養開始日と注されているが、事後MTA細胞リストの5月25日なら35日継代しているので11回も継代していることになる。増殖率実験をしているのではありませんから3継代もしたら樹立としてキメラ実験に使用するのが普通ではないか。しかも桂報告書はリストでは4月19日培養開始として本文中では5月作製としか書いてない。これだと5月1日であった事にもなりうる。意図的にごまかしているんですね。明確に共犯だということが分かりますね。事後MTAは木星さんが取り寄せて分かったので、これが無かったら国民は騙されたままであったでしょうね。
コントロールのES細胞キメラのジャームライントランスミッション実験は実際には行われていないと思いますね。若山さんのデータ捏造です。それを何とかごまかそうとして桂報告書は細胞リストに4月19日培養開始という嘘のデータを書き込んだ。それでもまだ間に合ってはいませんが、他方で4月はネイチャー誌への投稿(投稿日不明)とヴァカンティ氏の米国特許仮申請(4/24)があった。
最初のネイチャー論文にはジャームライントランスミッション実験は終わっていませんから書き込まれていません。そもそも小保方酸浴細胞核使用ntESの生殖能力確認ですから、リクルートのために書かせている論文とは無関係だったんです。4月のネイチャーリジェクトで後はヴァカンティ氏が自分の主催誌であるティシュー誌に掲載し、小保方さんを解放して自分に渡してくれるという話になってた筈だったわけです。ところがヴァカンティ氏がセルだサイエンスだと言い始めたのです。
ですから若山さんがネイチャーへの再提出に向けて<ES細胞の混入による可能性を否定するために、「ES細胞は同時に培養していなかった」という記事を原稿に加えることも指示>したのは別に嘘ではないわけです。まだ4月の論文提出時には129B6F1ES1~6は存在してなかった。小保方さんにそう言ってますから、逆に4/19より前にはこの作成時期を遡らせることが出来なかったんでしょうね。これは前提通りに作製していくと、キメラ胚へのインジェクションが4月20日前後ならその時にはES細胞は出来ていることになって、ES細胞は5日胚のインナーセルマスを取り出して培養を3継代程度確認したものでしょうからF1マウスの交配から12日程度かかっている。4月8日には計画しているということになる。私はESのジャームライントランスミッションなんて分かり切っていることなので若山さんがこんな実験をするわけがないと思いますね。予算の無駄です。ES細胞の増殖率実験も同じですね。やるわけがない。小保方さんにやらせるわけもないということです。対して小保方酸浴細胞核使用ntES細胞に関してはすべて確認しないといけません。未経験の細胞です。
第一レフェリーを誰それだと悟って若山さんがコンタクトをとっていたという可能性も考えられるんですが今はその問題は置いておきましょう。結構ややこしい。
それよりもこのレフェリーは刺激惹起の論旨を頭から否定しているので桃子が論調が変わったと言っているのは、二度目の提出で、しかも理研と笹井さんの名がありますから世辞も述べますね。前回のはヴァカンティ氏の名前です。
内容は厳しいものですね。これを若山さんが<三大誌3つのレビューワーコメントの中で、ネイチャーからのコメントが一番やさしいので>と言うところに何か第一レフェリーとの関係を疑義するわけです。
続きです。

>
>研究と結論は意図的にある特定の分野に焦点が当てられていて、他は除かれている。本文から読者は直ぐに刺激が多能性に向けた何らかのリプログラミングを引き起こしていると思ってしまう。このことはテラトーマとキメラ形成によって際立って提示されている。しかし、リプログラミングは一つの説明であるにすぎない。再生、脱分化及び増殖能力、そして異なる組織への分化は又腫瘍を含む癌幹細胞の特徴でもある。私は著者らに遺伝子操作なしの幹細胞様の作成法の発見に関する主張に注意深くあることをお奨めしたい。著者らは処置が遺伝子に引き起こしている実際の効果を調べ、遺伝子の不安定性に気づかないといけない。
論旨の全否定だということはアーティクル論文を読んでいる人には自明ですね。刺激惹起というのは手記では笹井さんが命名したと花を持たせた書き方になってますが、SACsという言葉は例の実験ノートにも以下のように出ている。
| 2月27日 |
| CD45+カルスのテラトーマ切る |
| Ac or rsc or sacs・・・ |
| AFP |
| βⅢ 1:200 |
| αSM |
無論サイエンス誌投稿論文ではStress Altered Cellsという命名で論じられていて、刺激惹起という発想そのものはもともと小保方さんが抱いていた仮説ですが、これを第一レフェリーはアーティクル論文においても全否定しているということです。
本文です。
>
>著者らは、体細胞系譜決定が遺伝子修飾に働きかけることによってのみ強力に変化させ得ると、又、その過程は自然には生じないと、読者を信じるように導いている。遺憾ながら私は同意しない。これは心筋細胞の部分的分化が心臓疾患治癒の原因となって以降のケースとは違う(Senyo et al.,2012 nature 事例を参照)。面白いことに、細胞系譜決定の動的平衡が癌幹細胞培養維持にも又示されてきている(Gupte et al.,2011 Cell 事例)。私見では、これらの事例は、著者らによって為されている主張に反して、哺乳動物体細胞は遺伝子修飾無しに、脱分化能力を持つことを示している。
ここはど素人としてはどちらが正しいのかを判断することはできませんね。笹井さんはただ英語と論文の書き方の指導をしてやって、この論文を通してやってくれと頼まれている。個々の実験に虚偽があるかもしれないと考えては出来ない仕事を頼まれている。一科学者としてとか、CDBのGD、つまり管理職としてどうあるべきかという判断は結構むつかしい判断で、笹井さんほどの人であるとこの論文はかなり実験の詰めが甘いということはすぐに感じたと思いますが、ただし、キメラがスタンダードな意味で書かれている通りに出来たということは疑えません。そもそも論文を通してやってくれと頼まれたことそのこともキメラができているという絶対的事実に基づているわけですから、ここを疑えということは、若山さんを疑うということであり、理研のシニアな学者を疑うということですから、常識的にまずは不可能です。そういう人なら理研に10年間も居た事自体の管理責任がどうなのだということになる。10年間そんなことは露なかったから10年満期勤務したので何かいけないかと言われても非難のしようがないのではないでしようか。
では、小保方さんが不正をしてキメラが出来たのだとして、この場合関係者全員が小保方さんに騙されたということになるわけですが、そもそも若山さんがキメラが出来たと証言しているのだから、若山さんが小保方さんに騙されて、キメラを作ったのだとしたら、関係者全員が騙された責任は若山さんにあるわけです。これを聞いて面白いと思った西川さんや、竹市さんや、笹井さんや、丹羽さんや、野依さんが信じ込んだのは若山さんが出来たと言ったからで、西川さんや、竹市さんや、笹井さんや、丹羽さんや、野依さんが信じ込んだのを間違いだということはできないということをまず確認しておかなけければなりませんよね。
若山さんを騙した小保方さんが悪いという話はその後に論ずべきことですから、小保方さんが若山さんを騙したから、西川さんや、竹市さんや、笹井さんや、丹羽さんや、野依さんが騙されたというのは間違いですね。混同しないようにしないといけませんよね。それとこれは違いますね。
まずはすべては若山さんが悪いのです。
次に小保方さんが若山さんを騙したのが悪いというのも又おかしな話で、若山さんは小保方さんに騙されてはいけない人です。武田邦彦教授は最初にここを指摘していますよね。「僕は自分の学問分野で自分の学生に騙されたりはしないよ。騙されないくらいの圧倒的知識差があるから教授をやってる。」と。これを事件の話に置き換えると、「若山さんは自分の学問分野で小保方さんに騙されたりはしないよ。騙されないくらいの圧倒的知識差があるから教授をやってる。」
若山さんは山梨大の教授として、そこに小保方さんを助手で連れて行こうとしたのです。
武田教授の説明が正しいなら、若山さんは小保方さんに騙されたことなど一度もないはずですよね。むしろ上から見ていたはずですし、こんなことは別に学問分野でなくてもどんな職場でもそうですね。普通は上から下はミエミエですよね。それが普通でないのだったら、人間は成長しないものだということになってしまう。
騙したのなら若山さんが小保方さんを騙したということしか考えられませんよね。それが世間常識というものです。それに何か目下が悪いことしたらどやしつけて内々で処理して外に恥をさらすなんてことはしませんよね。ところが若山さんは内輪で話し合うこともせずに「騙されたあ」と世間に公言しているんです。おかしいと思わない一般人はいません。
従って、笹井さんが学問的にこの論文をおかしいと思わなかったことを非難することはできません。仮に笹井さんが第一査読者であったら同じことをコメントしたかもしれませんよね。なぜなら第一レフェリーはキメラが出来たという若山さんの言葉を信じなければならないような義務もなければ、若山さんと同じ職場でしかもGDとして管理者の立場にあった関係を若山さんとの間に持っているわけでもないからです。笹井さんだって、同じく関係ない立場だったらもっと厳しいコメントすらしていても不思議はないでしょうね。
でも笹井さんは同じ研究所に属している研究仲間です。その若山さんが例えば何か、嘘でも、とても馬が合わずに嫌いな人であったとしてすら、キメラが出来たと言っていることを疑うということはあり得ないことで、そもそもそんな考えではこの研究の仕事はできません。第一レフェリーでもこの論文を自分の仲間が書いていたらキメラができていたことは信じざるを得ないのですから、論文を信じるということになる。信じないときにはだから何かアーティファクトが無いかと訝ることになるわけです。
従って三誌論文とも皆が皆ESコンタミではないかと疑ったのです。ここでもレフェリーはコンタミを疑っている。しかし、真実はESコンタミキメラではなくて小保方酸浴細胞核使用ntESキメラだっただけですね。若山さんの最初の他愛ない嘘が最後の最後まで明かされないままになってしまっただけの事件でしたね。若山さんは最後まで論文がリジェクトされると信じていたんです。それまではすべてうまくリジェクトされてきていますからね。
若山さんが山梨大に転居して後の二報同時を主張しているのは、実は裏で、ネイチャーのこの第一レフェリーに本当のことを言ってないかとも疑義していますが、今は考察がややこしくなるので置いているところですね。
酸浴させて癌化している細胞は、エピジェネティクな遺伝子修飾がリプログラムされていなくても、<遺伝子修飾無しに、脱分化能力を持つ>と言われても、笹井さんはキメラができていると信じてますからね。この第一レフェリーは出来てないと思ってるんです。できていたと思った瞬間に<遺伝子修飾無しに、脱分化能力を持つ>なんて主張は関係なくなる。細胞の細かい解析がどうであれ、キメラができていたら多能性細胞であることは証明され終わっている。遺伝子修飾されたままにキメラができるなんてことはありませんからね。
本文です。
>
>施された細胞処置が遺伝子異常を引き起こす可能性に加えて、大量細胞死を引き起こしていることも明白である。著者らはその後の実験の流れに沿って、生き残った細胞に意識を向けて、図1に細胞数計測見積を示しているが、それは恐らく最善の実験ではない。著者らは細胞死の数量を実証し、かつ示し、そしてこれをOct4-GFP観測と同時にCD45陽性細胞を共染色することで実行しなければならない。これはPI/Annexin-Vを使ってできる簡単な実験である。
この図1はオリジナルでどうなっていたのかは分かりませんが、最終版の図のFigure1-dに結果しているのではないでしょうかね。
レジェンドは以下です。
d, GFP+ (green) and GFP− (yellow) cell populations (average cell numbers per visual field; ×10 objective lens). n = 25; error bars show average ± s.d.
d,GFP +(緑)、GFP-(黄色)の細胞集団(視野当たりの平均細胞数、×10対物レンズ)。 試行数= 25;エラーバーは、平均±のs.d.を表示。
PI/Annexin-Vというのは死細胞を染色してFACS選別する実験のようですが、これはアーティクル論文の海賊版翻訳を中断させていますが、再開すべきですね。
まずFACSの実験は以下の二つです。
Figure 1-c
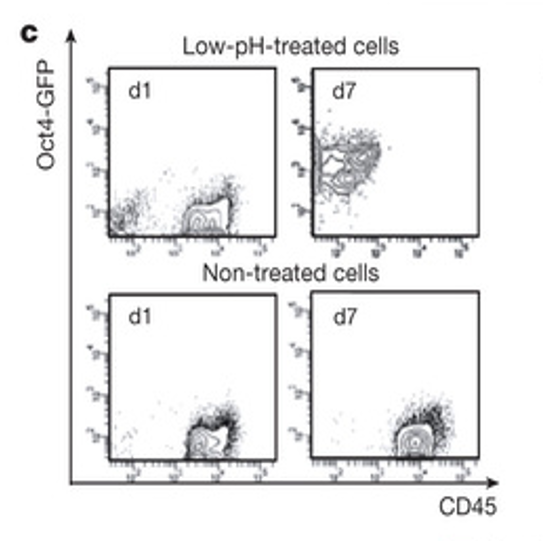
レジェンドは以下です。
c, FACS analysis. The x axis shows CD45 epifluorescence level; y axis shows Oct4-GFP level. Non-treated, cultured in the same medium but not treated with low pH.
c,FACS解析。 x軸はCD45の落射蛍光レベルを示す:y軸はOct4-GFPのレベルを示す。非処理(細胞)は同一培地であるが、低pH処理していない。
Extended Data Figure 1-b
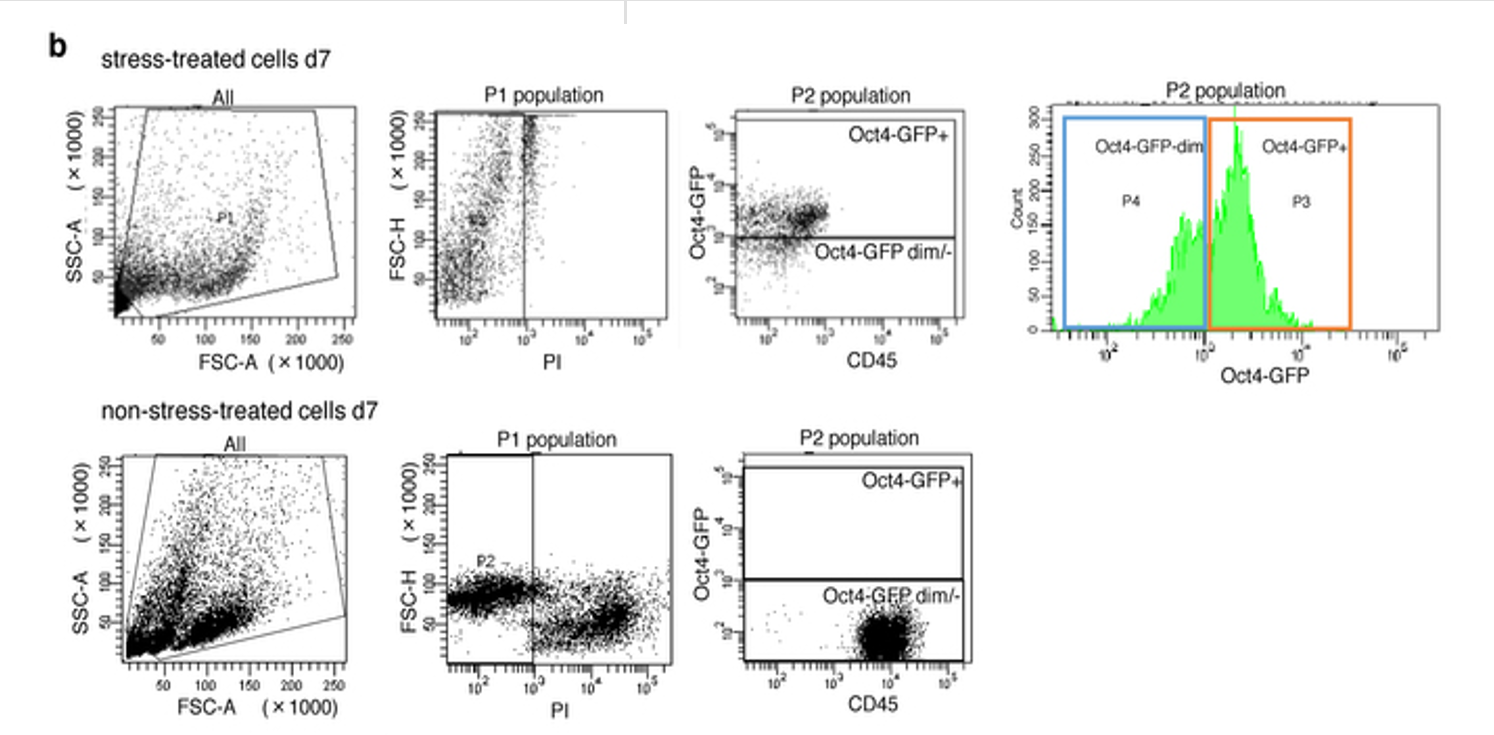
レジェンドは以下です。
b, Gating strategy for Oct4-GFP+ cell sorting. Top: representative results 7 days after the stress treatment. Bottom: non-treated control. P3 populations were sorted and counted as Oct4-GFP+ cells for all experiments.
b、Oct4-GFP陽性細胞ソーティングのゲーティング計画。上:ストレス治療から7日後の代表的な結果。下:未処理のコントロール。 P3集団を分類し、すべての実験でOct4-GFP陽性細胞としてカウントした。
そしてPI/Annexin-Vの実験は以下のExtended Data Figure 1-g,hです。
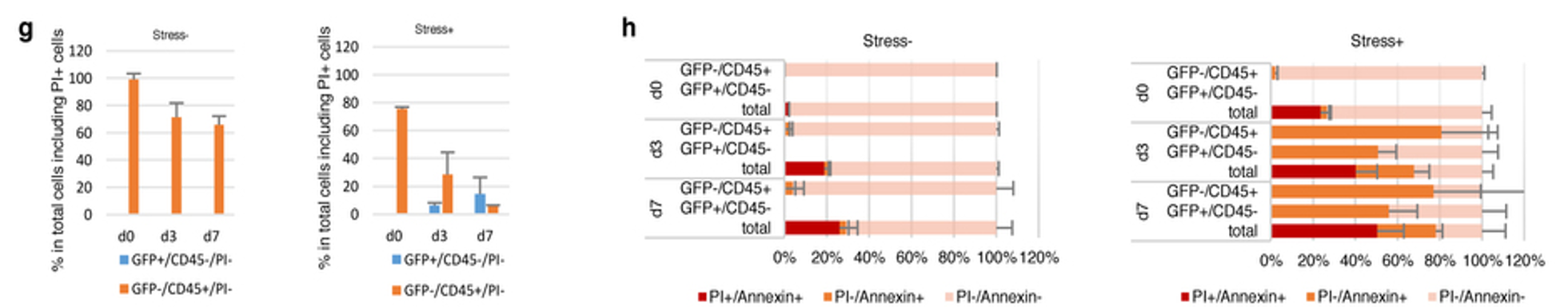
レジェンドは以下です。
g, h, Time course of apoptosis after the low-pH exposure. Stress-treated cells and non-stress-treated control cells were stained with CD45, annexin-V and propidium iodide at day 0 (immediately after stress treatment), day 3 and day 7. g, Blue bars, GFP+CD45−; orange bars, GFP−CD45+. Percentages in total cells included propidium-iodide-positive cells. h, Annexin-V-positive cells in these cell populations were analysed by FACS.
g、h、低pH曝露後のアポトーシスの時間経過。ストレス処理細胞および非コントロールの非ストレス処理細胞は、0日目(ストレス処理直後)、3日目および7日目に、CD45、アネキシンVおよびヨウ化プロピジウムで染色された。g、青色バー、GFP陽性CD45陰性。オレンジ色バー、GFP陰性CD45陽性。全細胞のパーセンテージにはヨウ化プロピジウム陽性細胞が含まれている。 h、これらの細胞集団におけるアネキシンV陽性細胞をFACSによって分析した。
まずFigure 1-cに関しては<海賊版翻訳アーティクル1>で以下のように考察した。
>>
**********
FACS( Fluorescence activated cell sorter)は蛍光活性化セルソーティングと訳されている。細胞にレーザー光を当てて識別後、電荷を与えて弾き飛ばす原理で分類できるらしい。
蛍光顕微鏡(fluorescence microscope)は強い励起光と弱い蛍光をフィルターを使って分離しているが、epifluorescence microscope というのがあって落射蛍光顕微鏡と呼ばれているらしいが、励起光を顕微鏡の中を通して照射するシステムで、ミラーをうまく使って蛍光のみを見たり撮影したりできる装置のようです。これはこれで専門的な知識が必要なようで、小保方さんは使い方を間違ってるとか言われていたりもしているが、ど素人には判断がつきませんね。
ただ、この画像はそんなに難しい操作結果だとも思えませんけどね。CD45+細胞は7日間変化がなく、だいたい同じ場所に飛ばされて集まっているが、酸浴細胞は1日目はたんなるCD45+細胞なのに7日目の細胞はGFP識別でほとんどが別の位置に飛ばされているということでしょう。
検出器の前を通るごとに識別されて分類される器械のようですね。こういう物理工学分野は素人にもすごいと分かりやすいが細胞はへなへなとしていて分かりませんねえ。
epifluorescence levelという意味は、光度指標か何かがあるのかどうかは分かりません。CD45+は何か抗体で蛍光染色しているはずですし、Oct4-GFPは発現すれば励起光を当てて確認するのでしょうが、両軸共にその光度か何かの目盛りにどんな意味があるのかは分かりませんね。10の何乗かという表示らしい目盛りの数値もよく見えませんが、ルクス表示のようですね。照度のようですね。
丹羽さんのGFPの漏れ出しは最大10%でした。この図はほぼすべてGFP発現していることになってますよね。蛍光しているスフィア塊だけを分析してますよね。因みに、言うまでもありませんが自家蛍光細胞はGFPでは選別されませんよね。自家蛍光だなんて言ってた人たちのお里の知れるところです。腹に一物あるんですね。ただ、その一物の道徳的評価はまだそんな判断の出来る段階ではありませんね。
**********
結論的には酸浴未処理細胞と酸浴細胞の酸浴直後のDAY0ではすべてがCD45陽性細胞(白血球)であるのに対して、酸浴DAY7では全部がOct4-GFP蛍光細胞になっているということです。
ただ、これは素人なので分からないのですが、これはCD45陽性でかつGFP陽性であった時には理論的には対角線上の辺りに集まると思いますが、全体がCD45の発現が弱まって左に寄っている。それが白血球が多能性細胞にリプログラムされたからなのか、それとも酸浴で白血球表面のCD45抗体蛋白が壊れて、その代わりに丹羽さんの発見したGFPの漏れ出しを検知しているのかは研究しないと分かりませんよね。
しかし再現実験ではキメラは出来ませんでしたので、この細胞がES並みの多能性細胞であることはないのですから、何かのアーティファクトなんですよね。ですから、ES細胞で若山さんにキメラを作らせたような人がこんなに真面目に研究しているわけがないと知れるところです。
論文ではキメラは出来ていると信じていますから、この現象を正しく考え直すなんてことは出来ませんよね。丹羽さんの発見したGFPの漏れ出しに気づくためには、キメラができないという事実から出発しないと無理なんです。小保方さんも、笹井さんもキメラは出来ているという前提でこの現象を見ている。
キメラができているという前提ならこのGFP発現は多能性細胞への転換だと考える方が当たり前だということに気づかないといけないですよね。ですから、査読者の「遺憾ながら私は同意しない」という疑義は必ずキメラは出来てないという根本的疑義に基づくことになるのです。
Extended Data Figure 1-bも<海賊版翻訳アーティクル1>で考察していますので省略しますが、何しろど素人なので機械の原理に関して無知ですから誤解も多いでしょうね。
Figure1-dはまさに査読者が「著者らはその後の実験の流れに沿って、生き残った細胞に意識を向けて、図1に細胞数計測見積を示している」と言っていて、かつその前に「研究と結論は意図的にある特定の分野に焦点が当てられていて、他は除かれている。本文から読者は直ぐに刺激が多能性に向けた何らかのリプログラミングを引き起こしていると思ってしまう。このことはテラトーマとキメラ形成によって際立って提示されている。しかし、リプログラミングは一つの説明であるにすぎない。」というアーティクル論文の論旨全体に対する全否定の向けられたまさにその解釈ですね。
笹井・小保方論文ではこの緑の細胞がキメラになったのだと解釈されている。実際キメラになったんです。論文はキメラになったという前提で書かれているんです。リヴァイズ実験中に理研内部でキメラになってないなどという解釈はありませんから、査読者はその解釈は可能性の一つに過ぎなくて他の解釈もありうると批判しているわけですが、笹井・小保方さんにはそんな視点は無い。
査読者はキメラはESコンタミだと思っていますから、別の解釈もありうることになるわけですが、この査読書は採用を考えているので、リヴァイズしてくれと要請してきているものですから、当然キメラが出来たことを認めているんですね。三誌論文はキメラは出来てない筈という判断でリジェクトされています。最初のネイチャーの査読者であった#1さんもキメラは出来てない筈と判断してリジェクトしているわけです。
できていると判断していながらリジェクトする商業雑誌は無いと思いますね。雑誌は売るのが目的ですから、こんな大発見を事実なのに掲載しないということはないでしょうね。事実でなくて掲載すると今度は信用失墜して売れなくなりますから、用心深く判断するわけです。
#1さんは要請を受けて、このキメラはESコンタミですよと意見しているわけです。だからこそ若山さんはそのときにESはなかったと書いておきなさいと指示しているわけです。
ところが今回は理研と笹井さんや丹羽さんのネームヴァリューが加わって編集部が信用したわけですよね。リヴァイズさせるという条件でコメントを書かせていますからこの#1さんもキメラは出来てないでしょうとは書けないんですね。ですからキメラ以外に関していろんな指摘をしている。
ここではまだででてきていませんがレターの査読では論文掲載をお勧めしないと書いています、二報同時だから再投稿できたのですから、レターは掲載しないということはやはり全否定なんですね。
キメラは出来てないでしようとはリヴァイズと要請している中では言えないので、別の解釈があるという指摘になる。それが死細胞の方をちゃんと数えなさいという指摘で、小保方さんはその指摘に応えてExtended Data Figure 1-g,hの実験を行ったのです。
こういうことを検討し始めると、直ぐに忘れがちですが、キメラが出来ていたら他の細かな雑多な証明は実は何も要らないのだという具合に証明法にもランクがあるということです。キメラが出来たらそれは多能性細胞です。Oct4遺伝子が発現したら多能性細胞であるかもしれない。なぜなら多能性細胞であるES細胞はOct4遺伝子を発現するからです。でもES細胞はキメラ証明されていますが、ES細胞でないものがOct4遺伝子を発現していたら、多能性細胞であるかもしれないから、次はどうするかというとキメラを実際に作ってみるわけです。何故ならOct4遺伝子を発現しているがまだキメラ確認されていないからです。
#1さんがどれほどおかしなコメントをしているかは理解できますよね。キメラは出来ていると認めた形でコメントしているのに言ってる内容はキメラは出来てないと言ってるのと同じことになるんですよね。
PI/Annexin-Vで死細胞を確認しなさいよと指示しているのですが、それを確認したら何が分かるのかというと、蛍光細胞が多能性細胞であるという可能性だけではないことが分かるでしょと言ってる。解釈はいろいろありうるのだよと言ってるんですが、ではあなたはなぜリヴァイズ要請コメントをしているのだというと、本心は別として、キメラが出来たということを認めたからでしようということになる。キメラが出来たことを認めると、この細胞は多能性細胞だということを認めたことになる。キメラは最終証明ですからね。
するとキメラが出来たことによって蛍光細胞の中の少なくとも何かが多能性細胞であったことが証明されたことになって、沢山あった可能性の一つ以外は否定されたということになるわけです。#1さんは自己矛盾を犯しているわけです。本来ならそれなら私は査読者を降りますというべきでしょうね。さもなければntESだということには気づかないまでもESのコンタミだから私は認めないと正直に書くべきなんですね。編集者との間に議論があったでしょうね。レフェリーとして残ったから苦しい書き方になっているわけです。桃子本では最後まで了承しなかったので編集部の責任で#1さんの意見は無視されてアクセプトされたといいますね。
GFP+というのは多能性細胞であるという判断です。無論キメラができているという前提です。
CD45+というのはまだ体性細胞である白血球だという判断です。
PI+は死細胞で核が赤く染色される。
g図はブルーがGFP+/CD45-/PI-ですから生きている多能性細胞という判断。オレンジはGFP-/CD45+/PI-ですから生きている白血球です。左図は酸浴未処理細胞群で全部生きている白血球です。d0からd7へ幾分生きている細胞数が減ってきているということです。縦軸は<% in total cells including PI+ cells>ですから死細胞も含めてFACS解析して生きている細胞だけを%表示したということです。右図は酸浴細胞群で、GFP-/CD45+/PI-とGFP+/CD45-/PI-の両方ができて来ている。酸浴直後のD0で既に20%の細胞が死滅していて、d7に向って生きている白血球が更に死滅していくが、生きている多能性細胞であるブルーの棒グラフが徐々に伸びて来るという図です。
この実験は#1さんがやらせた実験です。「施された細胞処置が遺伝子異常を引き起こす可能性に加えて、大量細胞死を引き起こしていることも明白である。著者らはその後の実験の流れに沿って、生き残った細胞に意識を向けて、図1に細胞数計測見積を示しているが、それは恐らく最善の実験ではない。著者らは細胞死の数量を実証し、かつ示し、そしてこれをOct4-GFP観測と同時にCD45陽性細胞を共染色することで実行しなければならない。これはPI/Annexin-Vを使ってできる簡単な実験である。」と言いましたよね。その結果、このブルーの伸びは「著者らは、体細胞系譜決定が遺伝子修飾に働きかけることによってのみ強力に変化させ得ると、又、その過程は自然には生じないと、読者を信じるように導いている。遺憾ながら私は同意しない。これは心筋細胞の部分的分化が心臓疾患治癒の原因となって以降のケースとは違う(Senyo et al.,2012 nature 事例を参照)。面白いことに、細胞系譜決定の動的平衡が癌幹細胞培養維持にも又示されてきている(Gupte et al.,2011 Cell 事例)。私見では、これらの事例は、著者らによって為されている主張に反して、哺乳動物体細胞は遺伝子修飾無しに、脱分化能力を持つことを示している。」ということを証明したでしょうか。
これは#1氏の論理飛躍のしすぎではないでしょうか。
でも、キメラは出来なかったという現在の知見からこれはどういう解釈になるのかというと、小保方さんの酸浴細胞は大半死に絶えながらも生き残った細胞は丹羽さんの検証実験によれば内在性Oct4はほとんど発現しないのに、マーカーであるGFPだけが強く発現する細胞が残ってくるということが分かったわけです。と同時に、亜致死下の細胞がすべて微量の内在性Oct4を発現していることも小保方さんによって証明されていましたね。
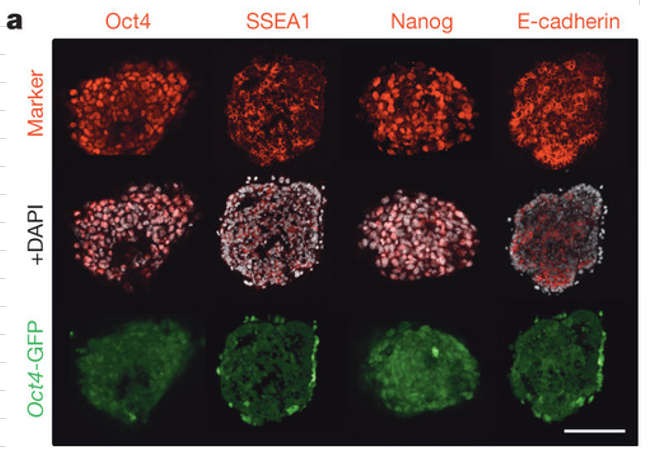
a, Immunostaining for pluripotent cell markers (red) in day 7 Oct4-GFP+ (green) clusters. DAPI, white. Scale bar, 50 μm.
a,7日目のOct4-GFP陽性(緑)クラスター内の多能性細胞マーカー(赤)の免疫染色。 DAPI、白。スケールバーは50μm。
左上です。これは免染結果ですから定性分析です。すべての細胞が微量に内在性のOct4蛋白質を発現しているんです。丹羽さんの漏れ出しの表は定量分析ですからGFP遺伝子の発現量に対しては内在性Oct4遺伝子はゼロになっている。
(a)Q-PCR analysis of the low-pH treated liver cells cultured for 7 days. Liver cells were prepared from 7-day old GOF mice and treated with either ATP or HCl, or without stressor. RNA samples were prepared from all cells in the wells at day 7 of culture and the relative expression levels of Gfp (derived from GOF Tg) and Oct3/4 (derived from the endogenous Pou5f1 allele) to Gapdh were indicated with standard deviation. The expression levels in control ES cells carrying CAG-GFP Tg were set at 1.0.
(a)7日間培養した低pH処理肝細胞のQ-PCR分析。肝細胞を7日齢のGOFマウスから調製し、ATPまたはHClのいずれか、またはストレッサーなしで処理した。培養7日目にウェル中の全細胞からRNA試料を調製し、Gfp(GOF Tg由来)およびOct3 / 4(内因性Pou5f1対立遺伝子由来)のGapdh<訳注:glyceraldehyde-3-phosphate dehydrogenase;GAPDH mRNAはさまざまな生理状態において発現量が変わらないと考えられている>に対する相対発現レベルを標準偏差で示した。 CAG-GFP Tgを保有するコントロールES細胞における発現レベルを1.0に設定した。
Oct4-GFPを仕込んだマウス由来のES細胞を使っていませんね。CAGマウスのES細胞です。その発現量をGapdhの発現量と比較してその比を1としている。対して酸浴細胞はOct4-GFPですからCAG-GFPとOct4-GFPの発現量は発現する時には同じ量発現するという前提になっているんですが、そのようなことは分かってないのではないですかね。一番いいのはOct4-GFPを持つマウスのES細胞をコントロールとして使うのがいいですよね。
しかし、焦点の問題として、Oct4-GFPが発現しているのに内在性Oct4遺伝子は発現してないということがあるのだという事実は証明されているわけです。ただ、その量はまずCAG-GFPの発現量が基準になっているのと、もう一つは相沢さんの指摘している酸浴細胞のGapdh値が動いているというところに信頼性が欠如しているわけですね。
ですから、小保方さんの酸浴細胞が何であるのかということはまだまだ解明されてはいないのです。でも、相沢さんが小保方さんと雑談中に聞き出したこれまでの経緯の話によって、キメラは若山さんの訳ありの方便の嘘だと分かってしまったので、急いですべてを有耶無耶にして終わらせたというだけの話です。論文通りのキメラではないのだと分かったのです。
次に、h図はまず赤色のPI+/Annexin+は完全死細胞で、オレンジ色のPI-/Annexin+は初期アポトーシス、ピンク色のPI-/Annexin-は正常細胞です。<GFP-/CD45+>は白血球細胞集団だと思いますが、<GFP+/CD45- Total>の意味がよく分からない。何と何のトータルなのか。特に左図の未処理細胞には<GFP+/CD45->はないことにg図でなっていますから、トータルしても同じはずですが、<GFP-/CD45+>自体より余計に細胞死している。ここは良く理解できません。
本文の続きです。
>
>著者らは観測されている過程が実際に細胞転換に思えるのか、腫瘍化なのかを定義するように努力しないといけない。比較はOct4-GFP陽性細胞と非処理CD45陽性細胞との間で行われているが、最善の内部コントロールとしては処理中にできてくるOct4-GFP陰性細胞の集団である。
非処理のCD45+細胞(白血球細胞集団)から酸浴処理後にOct4-GFP陽性細胞ができると同時に生きたままのOct4-GFP陰性細胞も残っているが、それはCD45陽性細胞(白血球細胞集団)ではないのかという疑義ですね。
本文は続きます。
>
>図1bは適正に示されていない。すべてのコントロール細胞を含む適切な識別手法が示されなければならない。同様に、それぞれのパネルはきちんとラベルしておかないといけない。読者はなぜ非処理のCD45陽性細胞が(三日目の)CD45の減少を示すことになったり、7日目までの完全に非処理の独立した細胞集団ですら減少することになるのか戸迷うことになる。又、著者らは、過剰造血細胞系列と前駆細胞を含むCD45陽性細胞としての出発点の細胞集団の特性定義を求められている。
どうやらオリジナルの図1bは最終版の図1dのようですね。
d, GFP+ (green) and GFP− (yellow) cell populations (average cell numbers per visual field; ×10 objective lens). n = 25; error bars show average ± s.d.
d,GFP +(緑)、GFP-(黄色)の細胞集団(視野当たりの平均細胞数、×10対物レンズ)。 試行数= 25;エラーバーは、平均±のs.d.を表示。
D0というのは酸浴直後ですから、ここには酸浴前のデータがないんですね。そもそも酸浴前のCD45+細胞の定義を求めているわけで、その後に今度は酸浴以降に常に幾分生き残っている、ここではOct4-GFP-とされている細胞集団の定義を求めている。つまりそれらはGFP発現がないのはいいとして、そもそもまだCD45抗原を持って維持しているのかということですね。最終図は以下です。
オリジナルでは以下のaがあって後に今のdがbとして提示されていたのでしょうかね。最終形ではその間に今のb,cが挿入された。
☟
☟
☟
この部分に関する本文は以下ですね。
Low pH triggers fate conversion in somatic cells
CD45+ cells were sorted by fluorescence-activated cell sorting (FACS) from the lymphocyte fraction of postnatal spleens (1-week old) of C57BL/6 mice carrying an Oct4-gfp transgene, and were exposed to various types of strong, transient, physical and chemical stimuli (described below). We examined these cells for activation of the Oct4 promoter after culture for several days in suspension using DMEM/F12 medium supplemented with leukaemia inhibitory factor (LIF) and B27 (hereafter called LIF+B27 medium).
Among the various perturbations, we were particularly interested in low-pH perturbations for two reasons. First, as shown below, low-pH treatment turned out to be most effective for the induction of Oct4. Second, classical experimental embryology has shown that a transient low-pH treatment under ‘sublethal’ conditions can alter the differentiation status of tissues. Spontaneous neural conversion from salamander animal caps by soaking the tissues in citrate-based acidic medium below pH?6.0 has been demonstrated previously.
Without exposure to the stimuli, none of the cells sorted with CD45 expressed Oct4-GFP regardless of the culture period in LIF+B27 medium. In contrast, a 30-min treatment with low-pH medium (25-min incubation followed by 5-min centrifugation; Fig. 1a; the most effective range was pH 5.4–5.8; Extended Data Fig. 1a) caused the emergence of substantial numbers of spherical clusters that expressed Oct4-GFP in day-7 culture (Fig. 1b). Substantial numbers of GFP+ cells appeared in all cases performed with neonatal splenic cells (n = 30 experiments).
The emergence of Oct4-GFP+ cells at the expense of CD45+ cells was also observed by flow cytometry (Fig. 1c, top, and Extended Data Fig. 1b, c). We next fractionated CD45+ cells into populations positive and negative for CD90 (T cells), CD19 (B cells) and CD34 (haematopoietic progenitors), and subjected them to low-pH treatment. Cells of these fractions, including T and B cells, generated Oct4-GFP+ cells at an efficacy comparable to unfractionated CD45+ cells (25–50% of surviving cells on day 7), except for CD34+ haematopoietic progenitors, which rarely produced Oct4-GFP+ cells (<2%; Extended Data Fig. 1d).
低ペーハー刺激による体細胞の運命転換
CD45+細胞は蛍光活性化細胞選別器(FACS)によって、Oct4-GFP導入遺伝子を保有するC57BL/6マウス(1週齢)の出生後の脾臓のリンパ球画分から選別された。そして様々なタイプの強い一時的な、物理的、化学的刺激に曝された(下記参照)。我々は白血病抑制因子(LIF)及びB27(以下LIF+ B27培地と呼ぶ)を補充したDMEM/ F12培地を使用した懸濁液中で数日間培養後Oct4プロモーターの活性化についてこれらの細胞を調べた。
さまざまな撹乱の中で、我々は2つの理由から低pHによる撹乱に特に興味を持った。以下に示すように、まず、低pH処理は、Oct4の誘導のために最も効果的であることが判明した。次に、古典的実験発生学は「亜致死」条件下での一過性低pH処理が組織の分化状態を変化させることができることを示している。 pH6.0以下のクエン酸ベース酸性媒体中に組織を浸すことによってサンショウウオのアニマルキャップからの自発的神経変換が以前に実証されている。
刺激に曝さないままだと、CD45でソートされた細胞はどれもLIF+ B27培地での培養期間にも関わらず、Oct4-GFPを発現しなかった。対照的に、低pH培地での30分間処理は(25分間の保温期間に5分間遠心分離<図1a>、最も効果的な範囲はpH5.4から5.8であった<拡張データ図1a>。)7日の培養においてOct4-GFPを発現する多くの数のスフィア状クラスターの出現を引き起こした(図1b)。 GFP 陽性細胞の大量の数は新生児の脾臓細胞で実行されたすべてのケースで現れた(30回の実験)。
CD45陽性細胞を使ったOct4-GFP陽性細胞の出現はまたフローサイトメトリー装置によっても観察された(図1c、最上部、及び拡張データ図1b、c)。我々は次にCD45陽性細胞をCD90(T細胞)、CD19(B細胞)およびCD34(造血前駆細胞)の陽性と陰性の集団に分画し、それらを低pH処理に供した。T細胞とB細胞を含むこれらの分画細胞は、Oct4-GFP陽性細胞をめったに作らなかったCD34陽性造血前駆細胞を除いて(2%以下;拡張データ図1d)、未分画CD45陽性細胞(7日目に生存している細胞は25〜50%)に匹敵する効果でOct4-GFP陽性細胞を発生した。
最後のところに脾臓から取得した白血球特異的分化抗原であるCD45を発現している細胞集団を更に分画して、CD90(T細胞)、CD19(B細胞)およびCD34(造血前駆細胞)とそれ以外のつまりCD45細胞の4つに分類したが、造血前駆細胞はGFPを発現する細胞にならなかったと書いている。
これは実際に小保方さんが#1さんに言われて行った実験ですが、これは大きな確認で造血前駆細胞は幹細胞からT細胞やB細胞に分化する直前の状態ですから幾分まだ多能性を持っている可能性がある。それはOct4-GFPを発現しないと確認しているわけです。もっとも前駆細胞をキメラ胚に入れてもキメラは出来てきません。これらの体性幹細胞はすでに全能性は失ってるんですね。ですからこの結果は別に小保方さんが強弁のために嘘をついているわけでもないんですね。単なる事実なんですね。
でも、結果的には小保方さんの細胞は論文通りのキメラなんてできてませんから、大本の嘘に騙されているわけです。でもそれは嘘だったのだとすると小保方さんのこの確認は何を意味しているかというと、GOFマウスに仕込まれているOct4-GFP人工遺伝子構造は他の細胞では丹羽さんの発見したGFPの遺伝子異常発現を引き起こす設計になってしまっているが、前駆細胞ではその異常は起こらないのだという類の一つの事実の発見になるんですね。するとOhoboさんたちが何故細胞を虐めると自分たちの設計した人工遺伝子が正常に発現しなくなるのかを突き止めるための重要なデータになりうるんですね。基礎研究というのはそういうもので、知らなかった事実をとにかくたくさん集めることで、それらが何かの時に新たな大きな発見に繋がることが千三つあるんですね。だからこういう事件になりましたが失敗なんて研究の常態ですから、本来は世間がとやかく言うようなことではないのです。
(2022/6/16)
酸浴細胞でなぜ内在性のOct4遺伝子は発現しないのにOct4-GFP人工遺伝子が発現するのかという問題は丹羽さんによってポンと投げ捨てられたのち関係者には放置されたままになっている。その理由は、これが事件の収拾策の中にあるからなんですね。事件が無かったとしたら、どうしてそんなことが起きるのだろうと興味深く研究者は思って、いろんな研究をした結果、論文が出るのが当たり前ですね。でも出てません。事件になってますから出たら世間が騒ぐから分かる。だから実際、出てないんですね。また、事件になってなければ論文は出たでしょうが、世間は気づきもしなかったでしょう。
要するにこの分野の研究界のタブーになってしまっているわけです。事件を蒸し返さないためですね。他にやることはいくらでもありますからね。それは誰にでも理解可能ではないか。
遺伝子というのは開始コドンがあって終始コドンで読み取りが終わります。で、読み取りの開始はどういう仕組みで起きるのかというと、構造遺伝子の前方にプロモーター領域という配列があってその部分に他の要因で作られた蛋白質が結合することが契機になって、いろんな種類があるが、代表的なものとしては、それまで読み取りができないようになっていた部分が取り外されることで読み取りが開始される。
GOFマウスには普通の遺伝子だけでなくGFP人工遺伝子が組み込まれている。
①[Oct4遺伝子プロモーター領域配列]+[Oct4構造遺伝子]---(内在性遺伝子)
②[Oct4遺伝子プロモーター領域配列]+[GFP構造遺伝子]---(人工遺伝子)
常識的に考えると[Oct4遺伝子プロモーター領域配列]に働きかける蛋白質が核内に入ると当然①も②も同時に発現しますよね。②の蛍光発現は①の発現を保証している。だからこそその便利さからGOFマウスとして販売されているんです。この蛍光遺伝子発現が無かったら免染で確認しないといけないですがマウスを殺さないと免染は出来ません。②があると生きているままで確認できるわけです。
ところが小保方さんが細胞を酸浴させるという乱暴な処置をしたら①は発現してないのに②だけ発現する現象があるという事実を丹羽さんが発見した。理屈上そんなことはあり得ませんよね。ならば理屈が間違っているんですね。誰が考えてもこんな単純な論理に間違いはありませんから、間違いは前提にあるに決まっている。
ど素人の推測では②の概念図に間違いがあるのだと思われますね。DNA配列はほとんどの部分がジャンク配列で無意味な配列が間にたくさんある。でも、それがどういう働きをしているのかはまだまだ解明途上なんですよね。分かってないんです。②は恐らく以下なんです。
②[Oct4遺伝子プロモーター領域配列]+<イントロン>+[GFP構造遺伝子]---(人工遺伝子)
①は例えば恐らく以下なんです。
①[Oct4遺伝子プロモーター領域配列]+<イントロン>+<イントロン>+[Oct4構造遺伝子]---(内在性遺伝子)
つまり人工遺伝子を作る時に切り取ってきた部分だけの働きと自然の配列には当然違いがあって、現在の知見ではそのイントロン部分には働きがないとされていて、実際に普通の状態では働かないが酸浴という特殊環境でイントロン部分に何らかの働きが生じて②だけが発現するというような可能性ですね。
すると、小保方さんが査読要請に従って前駆細胞ではGFP蛍光しないという事実を発見していることが、何か前駆細胞特異的な他の蛋白質の介在が漏れ出しを防いでいるのだと推定すると、①と②とのイントロン部分の違いの何がそうさせているのかの原因解明に寄与しうるデータとなるわけです。
ですから、常識で考えて、実験事実は事実として正確に報告しなければならないんですね。お金かかっていますから、事実を知るのに只ではないんですから、全人類が働いたお金の一部を研究に回して、ニュートン曰くところの浜辺の全砂粒の中の一つの砂粒程度に関して知るのに人類は何百年を使っているわけです。事実の集積があって初めて法則の構築が可能になる。具体的に何も分かってないのに法則が頭脳に浮かぶということはあり得ないので赤ん坊は何も知らないんですよね。
基礎研究って大変ですね。もっとも世間全員誰でも生きて行くのにはそれぞれ大変な思いをしてますけどね。
本文です。
>
>著者らはSTAP細胞が小さいということによって何を主張したいのか? 何の関係があるのか? 何故? 葉状仮足って何? 著者らは、これらの実験の背後にある疑問や仮説が何に関するものなのか、又何が結論として引き出されてくるのかを解説すべきだ。査読者である私にはこれらの観察結果の関連が示されているとは見えない。代わりに、クロマチン沈降実験と脱メチル化の情報提供をすると確かにより興味深いのではないか。必ずしもゲノムワイドプロファイリングまでは必要ではないが、簡単なヒストン修飾確認、かつ又は、脱メチル化確認実験があるととても論文の質が高まるのでないか。
ちょっと私の英語力では誤訳しているかもしれませんね。最初の方はいいんですね。細胞が小さいというのは事実を指摘しているだけです。当初は酸浴しているので中の水分が出て痩せているのではないかとも思っていたかもしれませんね。手記には細胞膜の損傷が関係してないかとも考えていたりしていたようですね。私は単にリンパ球の大きさが他の細胞種とは違ってかなり小さい細胞だったからだとみています。後に、若山さんの嘘として私は大きさの違いは分かったはずだとして以下の図を示しましたが、この査読段階では若山さんが胚盤胞にインジェクトする時に大きさの違いは分かったはずだなどと小保方さんが考えるわけもありませんから、後の事件化後の推測とは無関係で只小さいという事実を書いているだけです。
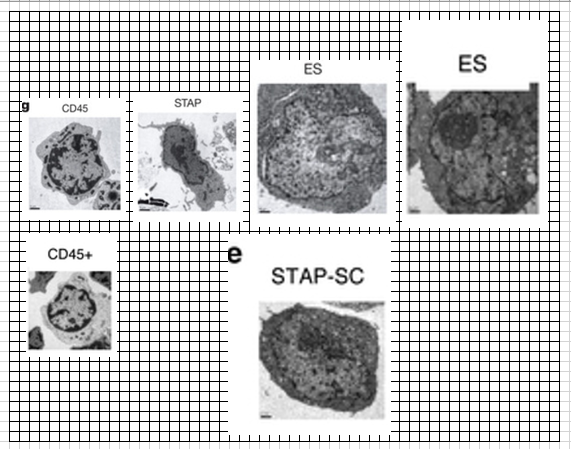
小保方さんはオリジナルの原稿にg図をつけていたのでしょうね。
査読者はこういうのを見て説明が足りないとクレームしているのでしょうね。
葉状仮足に関しては最終版の動画のレジェンドに以下のようにあるんですね。これは今でも見れます。本文は有料になっていますがサプリはダウンロードできます。私も以前これを見て笹井さんはここまでは見てやっていないのだろうなと思いましたね。リンパ球はアメーバではありませんから仮足運動はしませんので、笹井さんが見てやっていたら、これは焦点が合わせられていないから見えないが、マクロファージが死細胞を食べて動いているから引きずられているのだということはすぐに分かったはずです。小保方さんはリンパ球そのものが動いていると感じていたわけですが、深くも考えずに見たままに書いている。見たままの描写としては間違ってはいないわけです。
>>
| Live imaging of low-pH-treated CD45+cells |
|
DIC images during day 0 – day 7, overlaid
with oct3/4::GFP (green). A strong contrast of DIC (as compared to video
2) was applied to imaging so that lamellipodia-like processes (frequently
seen on and after day 4) could be viewed easily. (MOV 23217 kb)
|
低pH処理されたCD45陽性細胞のライブイメージング
0日目から7日目までのDIC[Digital Image Correlation:デジタル画像相関法]画像にoct3/4 :: GFP(緑)を重ねたもの。 DICの強いコントラスト(ビデオ2と比較して)がイメージングに適用されているため、葉状仮足のようなプロセス(4日目以降に頻繁に見られる)を簡単に見ることができた。 (MOV 23217 kb)
査読者にいろいろと言われてこの部分に関する最終稿の本文は以下になった。
>>
Low-pH-treated CD45+ cells, but not untreated cells, gradually turned on GFP signals over the first few days (Fig. 1e, Supplementary Videos 1 and 2 and Extended Data Fig. 2a), whereas CD45 immunoreactivity became gradually reduced in the cells that demonstrated Oct4-GFP expression (Fig. 1f and Extended Data Fig. 2b). By day 5, the Oct4-GFP+ cells attached together and formed clusters by accretion. These GFP+ clusters (but not GFP− cells) were quite mobile and often showed cell processes on moving (Supplementary Video 1).
The Oct4-GFP+ cells demonstrated a characteristic small cell size with little cytoplasm and also showed a distinct fine structure of the nucleus compared with that of parental CD45+ lymphocytes (Fig. 1g). The Oct4-GFP+ cells on day 7 were smaller than non-treated CD45+ cells (Fig. 1g, h and Extended Data Fig. 2c) and embryonic stem (ES) cells (Fig. 1h), both of which are generally considered to be small in size. The diameter of low-pH-treated CD45+ cells became reduced during the first 2 days, even before they started Oct4-GFP expression (Fig. 1f), whereas the onset of GFP expression was not accompanied by cell divisions. Consistent with this, no substantial 5-ethynyl-2′-deoxyuridine (EdU) uptake was observed in the Oct4-GFP+ cells after the stressor (Extended Data Fig. 2d).
CD45の免疫反応性がOct4-GFP発現の証明された細胞の中で徐々に減少したのに対して(図1f及び拡張データ図2b)、未処理細胞で無い低pH処理されたCD45陽性細胞は最初の数日間で徐々にGFPシグナルを発し始めた(図1e、補足動画1、2と拡張データ図2a)。5日目では、Oct4-GFP陽性細胞が互いに密着し、集積によってクラスタを形成した。これらのGFP陽性クラスター(ただし、GFP陰性細胞ではない)は大変可動で、しばしば動いている細胞過程を示した(補足ビデオ1)。
Oct4-GFP陽性細胞は細胞質のほとんどない特徴的な小細胞サイズを示し、また親のCD45陽性リンパ球のそれと比較して異なる核の微細構造を示した(図1g)。7日目のOct4-GFP陽性細胞は非処理CD45陽性細胞と胚性幹(ES)細胞(図1h)よりも小さかった(図1g、hおよび拡張データ図2c)。どちらも一般的にサイズの小さい細胞であると考えられている。低pH処理したCD45陽性細胞の直径は、GFP発現の発生が細胞分裂を伴わなかったにも関わらず、それらがOct4-GFP発現を開始する前であってすら、最初の2日の間で小さくなりだした(図1f)。これと同調して、いかほどの5エチニル-2'-デオキシウリジン(EdU)の取り込みもそのストレス因子後のOct4-GFP陽性細胞の中に認められなかった(拡張データ図2d)。
サイズに関してはCD45+細胞とES細胞のどちらも一般的には小さい細胞だと書いていて、酸浴後のSTAP細胞は細胞質が減ってもっと小さいと強調していますね。でも私が指摘しているように、小保方さんはサイズバーを正確に合わせていません。これは三つを並列して見やすくするためで、他方でキメラができていますから安心しているのでしょうが、上述しているように実験は事実通りに報告しないといけません。サイズバーを合わせると私が作った図になる。
重大なのは実寸比較していたら、単にES細胞がCD45+細胞より大きいということだけでなく、STAP幹細胞がESの大きさになっていることに気づけたはずなんですね。若山さんがntES化しているからES の大きさになっている。
小保方さんは<The Oct4-GFP+ cells demonstrated a characteristic small cell size with little cytoplasm and also showed a distinct fine structure of the nucleus compared with that of parental CD45+ lymphocytes (Fig. 1g). The Oct4-GFP+ cells on day 7 were smaller than non-treated CD45+ cells (Fig. 1g, h and Extended Data Fig. 2c) and embryonic stem (ES) cells (Fig. 1h), both of which are generally considered to be small in size.>と書いてますよね。こうであろうという自分の仮説の言葉に引きずられているから事実が事実通りに見えてないんです。言葉の力というのは大きいんですね。一般にも「思い込み」といいますね。視覚という直覚は言語と強く結びついていて、見えているものを関連付けて理解しようと努力するんですが、その時に自覚がないと「思い込んだ」理解のために不都合なものが見えなくなるんですね。
私が誤訳かもしれないと思うのは以下の部分です。
>>
代わりに、クロマチン沈降実験と脱メチル化の情報提供をすると確かにより興味深いのではないか。必ずしもゲノムワイドプロファイリングである必要まではないが、簡単なヒストン修飾確認、かつ又は、脱メチル化確認実験があるととても論文の質が高まるのでないか。
以下のInstead以下です。前の疑問とどういう繋がりになっているのかよく分からない。

Insteadの意味が前述した疑問に関して、そんな比較をするよりもChIPシーケンスや脱メチル化確認をした方がいいという意味なら、これはちょっと変なんですね。オリジナル投稿論文にはメチル化実験が無かったということになるんですよね。でも、この実験は最初のネイチャー投稿時に添付されていたデータだと桂報告書に書かれていましたよね。スライドです。

本文です。
| 2)Article Fig.2c について |
| ・メチル化を示すいくつかの黒丸および白丸の整列に乱れがある点(Oct4-GFP+ cells の Oct4 promoter) |
| ・DNA メチル化解析データについて Oct4 の CD45+ と Cultured CD45+ 、および Nanog の ES と Cultured CD45+ 、Nanog の CD45+ と Cultured CD45+ が酷似している点 |
| ・オリジナルデータとの不一致がある点 |
| (調査結果) |
| CDB 若山研におけるプログレスレポート(PR)にて提示された資料、論文原稿の各バー ジョンで示された図、実験を担当した CDB 若山研メンバーより提供された実験ノート記 録、GRAS のコンピューターに残っていた実験データを照合し、PR 資料や論文図に示され たデータの信憑性を検討した。また、小保方氏に作図法やデータ処理について聞き取り 調査を行った。その結果、以下のことが判明した。 |
| (1)図として提示されている結果は、以下のような経緯をたどっていた。PR 資料では、 2011 年 9 月 22 日に最初のデータ(非処理細胞、スフェア Oct4 発現細胞、および ES 細胞のOct4遺伝子およびNanog遺伝子のプロモーター領域のメチル化)が提示され、 その後、2011 年 11 月頃にも提示された。この 2 回の資料は同じ実験結果を示してい ると判断されたが、これらデータの真贋性を裏付ける実験データやノート記録を確認 することはできなかった。なお、これら PR 資料で ES 細胞とスフェアの結果が入れ替 わるなど、小保方氏のデータ取扱いの杜撰さがうかがえた。 その後、2012 年 4 月 12 日付けの PR 資料に全く別の実験結果が図として提示され、 この図は 2012 年 4 月の Nature 投稿原稿、Cell 投稿原稿でも使われ、最終的に Article Fig.2c として発表された。この図では、メチル化を示す黒丸の配置に一部乱れがあ り、手動で作図したと考えられた。また、CD45 陽性細胞(CD45+ および Cultured CD45+ ) のデータについては、両者のパターンが酷似していた。 また、STAP 幹細胞についてもメチル化解析が行われ、この結果は 2013 年 3 月に投 稿された Letter 原稿 Fig.3 として初めて提示され、最終的には Article Extended Data Fig.8d として発表されている。この図でも CD45 陽性細胞におけるメチル化が 示されているが、Oct4 遺伝子、Nanog 遺伝子いずれのプロモーターも Article Fig.2c と比較して、メチル化程度が低いことが特徴的である。 |
| (2)CDB 若山研メンバーが GRAS に依頼し取得され、GRAS に残されていた配列データを解 析した結果、これら配列データを用いて Article Extended Data Fig.8d が作図されたと考 えられた。ただし、データの選別(シークエンスした DNA クローンの選別)が行われており、 また、STAP 幹細胞については異なる細胞のデータが作図に使われるなど、意図的なデータ 取扱いがあった可能性を否定できない。 |
| (3)Article Fig.2c は、2012 年 4 月に投稿論文原稿図として現れているが、それ以前 に小保方氏は計 3 セット(2011 年 10 月 27 日 1 セット、同 11 月 17 日に 2 セット)、 GRAS に DNA 配列解析を依頼していることが判明した。そのサンプル名には bisulfite とあること、また 11 月 17 日の 2 セット分についてはそれぞれ「oct4」、「nanog」と の記述もあることから、これはメチル化 DNA 解析であったと判断した。「oct4」につ いては 96 クローンのシークエンスが行われ、作図に利用可能な高精度配列情報は 74 クローン分であった。しかし、この 74 クローン分のデータを用いて Fig.2c の Oct4 プロモーターの図を作図することは不可能であった。例えば Fig.2c では 11 か所中メ チル化部位が 1 か所以下のクローンが 18 クローンあったことを示すが、シークエン ス結果でこのようなパターンを有したクローンは 3 クローンのみであった。アライメ ントできなかった低品質配列クローンを含めても、この図を作ることは不可能であっ たと考えられた。 また、「nanog」については 96 クローン中 40 クローンが作図に使用可能と考えら れたが、これらを用いても Fig,2c に示された Nanog プロモーターの結果は得られな い。Fig.2c には 100%メチル化クローンが 15 クローン存在するが、シークエンス結果 でこのようなクローンは最大で 7 クローンしか存在しなかった。アライメントできな かった低品質配列クローンを含めても、この図を作ることは不可能であった。 |
| (4)小保方氏の聞き取り調査から、メチル化のデータを取りまとめる際に、仮説を支 持するデータとするために意図的な DNA 配列の選択や大腸菌クローンの操作を行っ たことが確認された。この点について、小保方氏から誇れるデータではなく、責任を 感じているとの説明を受けた。 |
| (評価) |
| CDB 若山研の PR 資料において図の取り違えがあったこと、Article Fig.2c について裏 付ける実験記録の存在が確認できないことなど、小保方氏のデータ管理は杜撰であった。 のみならず、小保方氏は、自認するとおり、得られたデータのうちの一部だけを仮説に 沿って意図的に選別して提示し、データの誤った解釈を誘導する危険性を生じさせた。 小保方氏はこのような危険性について認識しながらデータを選別したうえ、手動で作図 して存在しないデータを新たに作り上げたものである。よって、捏造に当たる研究不正 と判断した。 このようなことが行われた背景には、共同研究者によるデータに対する過剰な期待が あったことが推察された。若山氏は、上記のメチル化解析を小保方氏が行った研究室の 主宰者であり、シニア研究者として小保方氏を指導監督するとともに、共同研究者とし て、データの正当性、正確性について十分な注意を払うことが求められていた。若山氏 はデータの意図的な選別・提示に直接的に関与したとまでは認められないが、小保方氏 が若山氏の過剰な期待に応えようとして捏造を行った面も否定できない。少なくとも若 山氏は、小保方氏の指導監督を怠り、データの正当性、正確性について検証することな く、このような捏造を誘発したと認められ、その責任は過失とはいえ極めて重大である。 |
私の査読書読み取りでは、サイエンスベースの12/25ヴァージョン原稿を笹井さんが書き直してやった時のオリジナル投稿原稿には、最初のネイチャー、セルにはあったこのメチル化実験結果は無かったという解釈になる。何か誤解誤訳しているかなあ。
桂報告書の書き方でも「この図は 2012 年 4 月の Nature 投稿原稿、Cell 投稿原稿でも使われ、最終的に Article Fig.2c として発表された。」としていてサイエンス原稿に関して触れてませんね。小保方さんは「これでは論文に使えないね」と言われてモチャモチャやったことを認めています。それは最初のネイチャー論文に掲載されて、セルまでは使われたが、小保方さんはサイエンスではもういいだろうと思って外していますよね。この時点で不正は無くなっていた。そしてサイエンスもリジェクトされたのでサイエンスベースでの改訂版を作製していて、それを理研に呼び戻された時に日本への飛行機の中で12/25と日付を打ったんですね。そしてそれを笹井さんが書き直すことになった。書き直しのときに論文原稿に存在してないものは二人で討論はできませんね。するとこのとき無かったものを付けさせる契機になったのは査読者のこのコメントだということになるのですが、この#1さんは最初のネイチャー論文の査読者ですから、そこにメチル化実験結果が添付されていたのを知っているんですよね。
他方で「STAP 幹細胞についてもメチル化解析が行われ、この結果は 2013 年 3 月に投 稿された Letter 原稿 Fig.3 として初めて提示され、最終的には Article Extended Data Fig.8d として発表されている。」と書かれていることによって、レター論文に関して笹井さんが書いた3月投稿時に既にメチル化結果が添付されていたとしているのが本当なら、アーティクル側のメチル化実験結果もつけられていたと考えるのが普通です。ただし、誤解のないようにしなければいけないのはSTAP幹細胞はntESですから脱メチル化しています。仮にES細胞でも脱メチル化している。捏造の必要はありません。対してSTAP細胞は小保方さんが自分で作った段階で行っていますから、脱メチル化してないんですね。でもキメラは出来たと思い込まされているのでモチャモチャやったんです。ESでchimera捏造なんてしてたらモチャモチャすることも無いと分かりますよね。
さあ、査読者はアーティクル論文に脱メチル化結果があったらいいのにと言ってるのなら、最初はついてなかったことになる。私の誤読なんですかね。
そもそもこれをつけた方がいいのは若山さんの都合だと考えるとこの査読者と通じていないかと私が最初に疑った理由も理解できると思います。
そしてでも、レターにはついていたようなんですよね。
小保方さんは酸浴細胞はモチャモチャやったので外したが、レターは事実通りだったから添付したのでしょうかね。
特許書類ではACCsになっていて、これはネイチャー、セルの呼び名です。サイエンスではSACsですね。
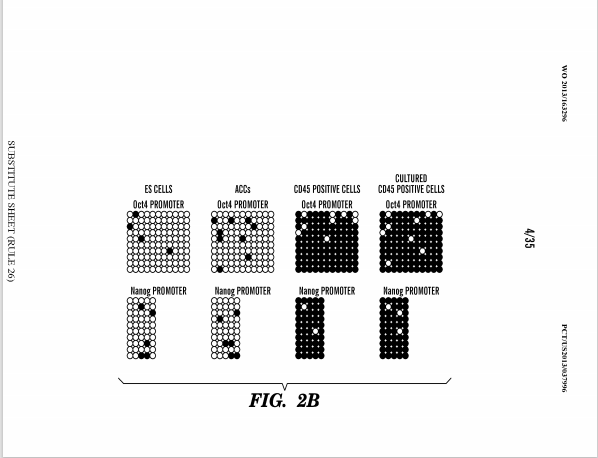
対してリトラクトされたレター論文にあった図はヴァカンティ氏は特許書類から現在全部外しているのですが、そこに「STAP 幹細胞についてもメチル化解析が行われ、この結果は 2013 年 3 月に投 稿された Letter 原稿 Fig.3 として初めて提示され、最終的には Article Extended Data Fig.8d として発表されている。」とされた同じ図がアーカイヴとして残されている。無論本文では抹消されている。
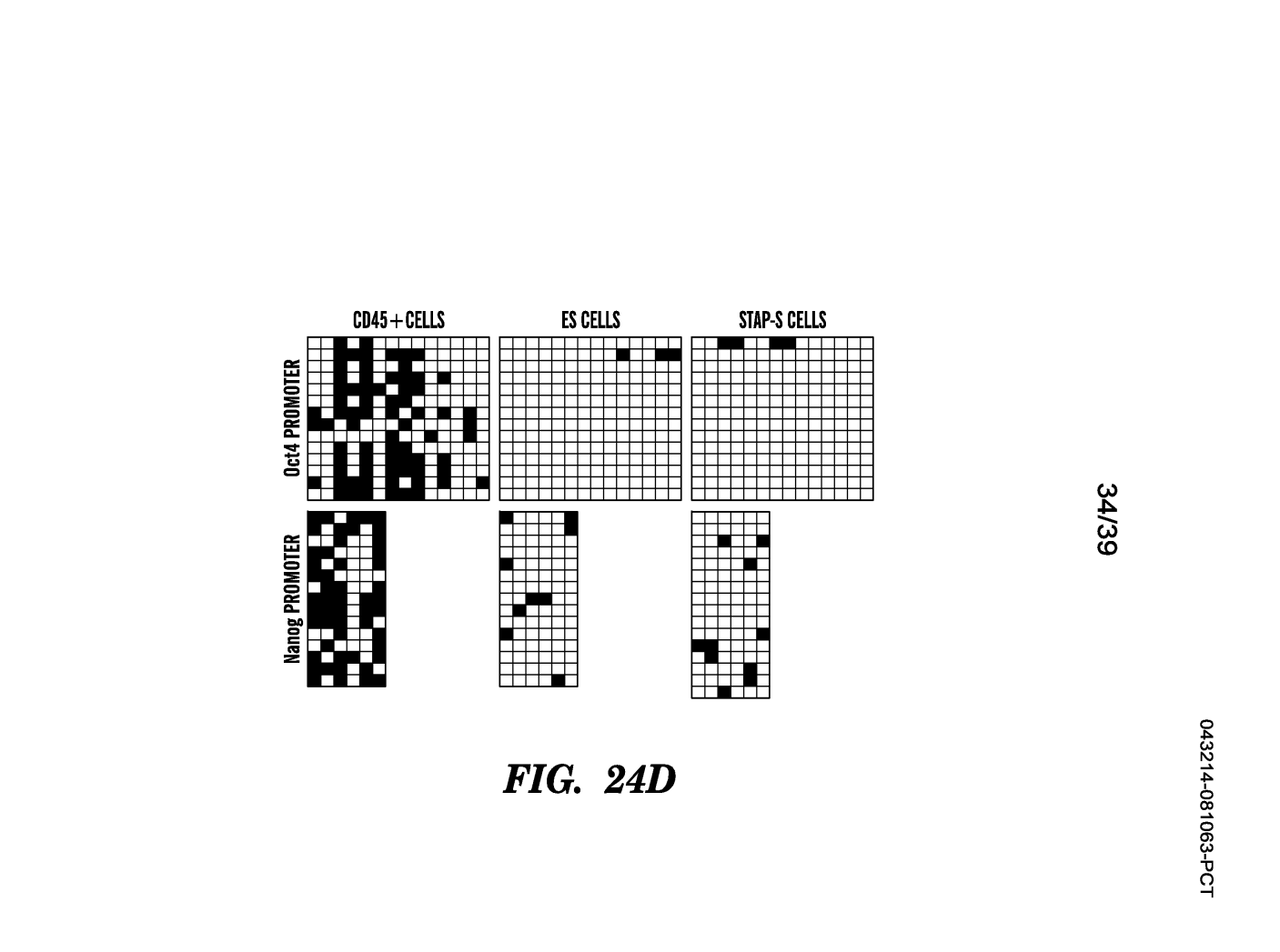
STAP-S CELLSとされているから削除されたんです。リトラクトされたネイチャー論文にあるものだから削除した。つまり三誌には無かったということで、これは三誌はキメラが出来たという論文で、幹細胞化論文は若山さんの研究ですからもともと無いんですね。
すると、このデータは若山さんが小保方さんに渡した幹細胞化のデータの中に含まれていたということになる。サイエンスベースの12/25ヴァージョン原稿の中にはもうメチル化実験の図は外されているが、若山さんの研究データの中には原稿はまだできてないがデータとして存在していたわけです。笹井さんはレター原稿を書いたときにそれを図として取り込んだ。しかし、アーティクル側に同じ分析がないことには気づかなかった。とすると、#1さんはレター原稿との比較はしなかったのかということになる。
無論若山さんのデータの中にあったとはいえ、実際に確認実験を行わされているのは小保方さんでしょうね。若山さんは酸浴細胞の脱メチル化実験を小保方さんにさせたと同時に、STAP幹細胞の脱メチル化実験も同時にやらせているんですよね。自分の酸浴細胞は脱メチル化してないのに、自分にはできないが若山さんの培養したSTAP幹細胞は脱メチル化していて、かつどちらもキメラが出来たと言われているわけです。その両方の結果を見て小保方さんは何故だろうと思わなかったのでしょうかね。
こういう問題の出し方というのはとてもテスト臭くないですかね。








コメント