



















臓器形成(発生・再生)は、情報伝達により臓器を構成する増殖・分化・運動により形態の変化を伴います。この形成機構を調べることは、すなわち情報伝達と形態変化を同時に研究することになり、両者の変化を時・空間的に正確に捉えることができるイメージング手法が重要となります。イメージングで"情報伝達を見る"、なおかつ"形の変化を見る"ことにより如何なるシグナルが臓器形成・形態形成を調節しているのかを解明したいと考えています。
循環器は心臓・血管が主な臓器となりますが、発生ではこの両臓器が他の臓器形成に先行して形成されます。他の臓器への酸素・栄養源の供給のためには、必須の臓器であるからです。心臓も血管も中胚葉から発生します。心臓原基となる細胞も血管内皮細胞となる細胞も中胚葉由来であり、両側中胚葉が正中に移動してきて原始心筒や初期大血管を形成します。さらに循環調節臓器となるべく両者がうまくcoordinateされた発生が進んで(心臓と大血管ー末梢血管が繋がる)いきます。肺で酸素化された血液を末梢までポンプとして送達し、抹消からの血液を心臓に戻すシステム(循環器)が完成します。従ってこの循環器の初期発生のメカニズムを調べることにより心臓や血管を新たに構築する(新生)ことに役立てたいと考えています。
メカニズムの解明には、分子生物学・細胞生物学的に解析することが必要であり、如何に臓器としての循環器が形成されていくのか?イメージング技術を駆使して調べていくことが目標達成につながると考え研究を行っています。われわれは、発生から学んだ臓器形成システムの理解を追及することで循環器再生にも貢献できればと思案しています。
※画像をクリックすると拡大します
(1) 心筋細胞の分裂のメカニズムの解明
(2) 心臓の左右非対称性を生む機構の究明
(1) 血管新生のシグナル伝達の解明
(2) 血管成熟機構の解明
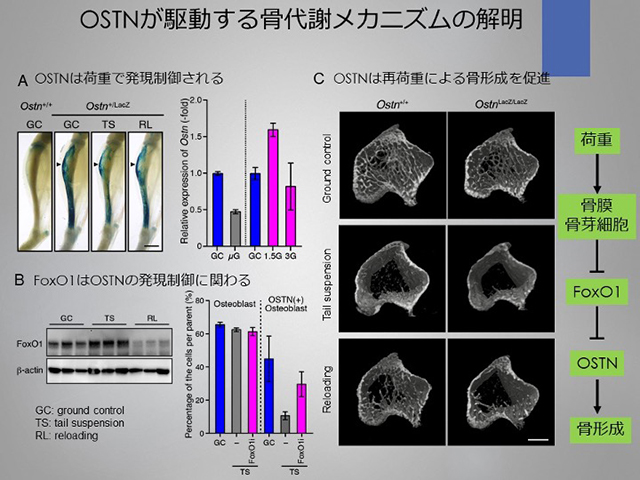
C.骨膜由来分泌性ペプチドホルモン
Osteocrin (OSTN)の生理作用
(1) マウス骨膜由来のOSTNの循環調節系への効果
哺乳類の心筋細胞は、出生後には分裂しないと考えられている。心筋細胞の分裂停止機構についても詳細な解明はなされていない。しかし、最近の研究では若年期までは2核になった細胞も細胞質分裂により、細胞数が増えることも報告された。一方、成体ゼブラフィッシュ心臓は障害があると既存の心筋が分裂を開始することにより、心臓が再生される。従って、ゼブラフィッシュ心筋細胞と哺乳類心筋細胞の分裂増殖メカニズムの相違を明らかにすることで心筋細胞の増殖を誘導できる可能性を検討したい。
細胞周期を可視化するFUCCIを心筋細胞特異的にゼブラフィッシュ心臓に発現させて、初期発生における心筋細胞増殖メカニズムを検討している。

心臓特異的にFUCCIを発現する
トランスジェニックフィッシュの心臓
緑:S/G2/M期の心筋細胞
赤:G0/G1期の心筋細胞

※画像をクリックすると拡大します
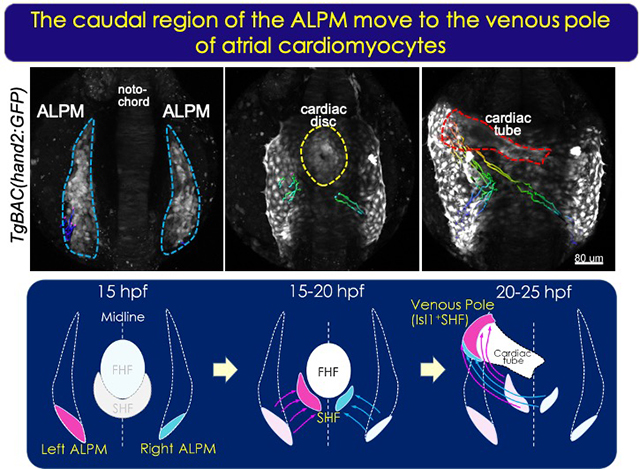
心臓は、初期発生では両側中胚葉からの心臓前駆細胞が正中線に向かい原始心筒を形成し、その後心房・心室・弁が形成されて同じ中胚葉由来の血管とつながる。心臓も、心先部が左に向き、大動脈も弓部となることで左右非対称となる。
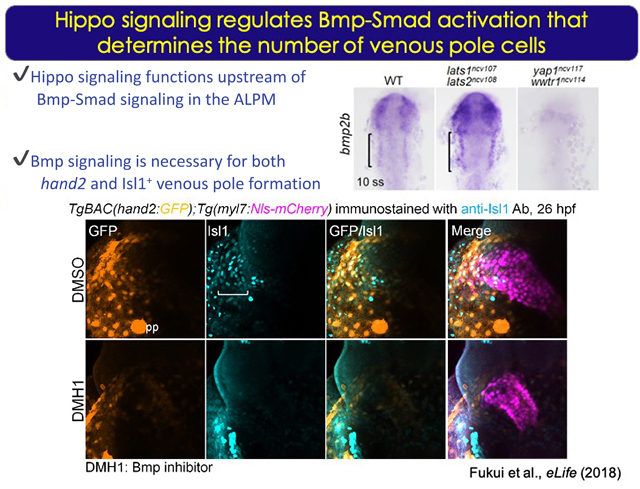
哺乳類では発生の初期に一時的に出現するノードによる左右非対称性の形成が、将来の臓器位置の決定に重要であり、ゼブラフィッシュでもノードに相当する Kupfferユs vesicle内のciliaによる流れが左右軸を決定する。我々は、このKupfferユs vesicleの形成過程のシグナル伝達の研究により、心臓の非対称性を理解したいと考えている。
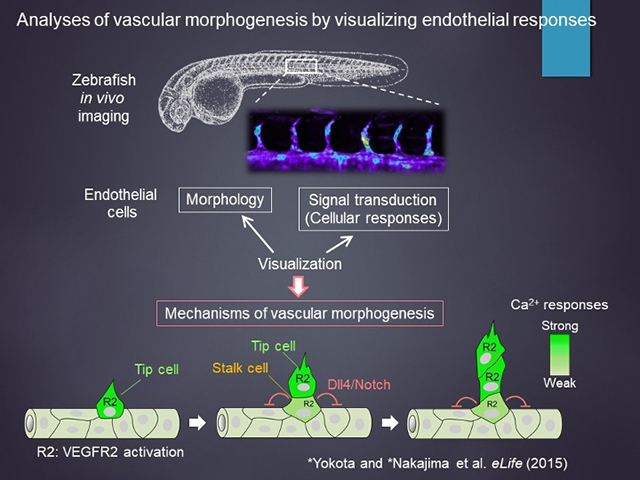
血管新生は、血管の発芽/分枝・内腔形成・剪定・成熟課程からなる。このすべての現象を調節する分子メカニズムを理解するためには、これらの生物学的現象を評価するシステム、すなわち、生きたまた見てそれが促進或いは抑制されているのかを判定する系の構築が不可欠と考えた。このためにゼブラフィッシュの背側大動脈から節間血管の形成を通じての血管新生課程をまず可視化して、上記のそれぞれを可視化できるトランスジェニックゼブラフィッシュを構築してきた。
まずは、血管形成の全過程を可視化することから始め、どのプロモーターがどの現象を捉えるのに至適であるのかを調べた。
※画像をクリックすると拡大します
そして、そのプロモーターを用いて
分枝・発芽:Rhoファミリー分子の活性化可視化
(1) 内腔形成: 極性可視化
(2) 剪定: アポトーシスあるいはオートファジー検出
(3) 伸長:細胞分裂
などを可視化できるトランスジェニックフィッシュを作製して解析を行っている。

ゼブラフィッシュの血管内皮細胞が、
大動脈から背側に伸張していく過程を可視化
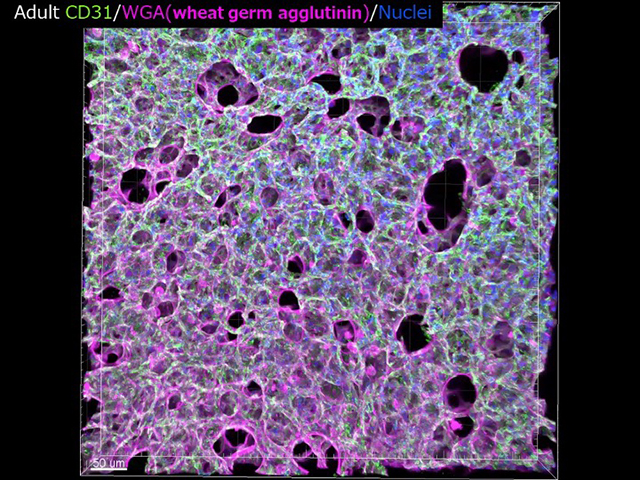
血管は一度構築されると内皮細胞障害(動脈硬化症)・虚血あるいは癌などの病態以外では、内皮細胞は安定した状態で維持されると考えられている。血管内皮細胞のturnoverは癌血管内皮細胞(数日)と比較して非常に長く100日から数年にも維持されると計算されている。(我々もゼブラフィッシュでFUCCIを血管内皮細胞で発現する個体で内皮細胞の発生における分裂を検討したところ、DLAVを形成してcaudal veinができあがる時期には、内皮細胞のほとんどG0/G1になってしまっていることが解った。)
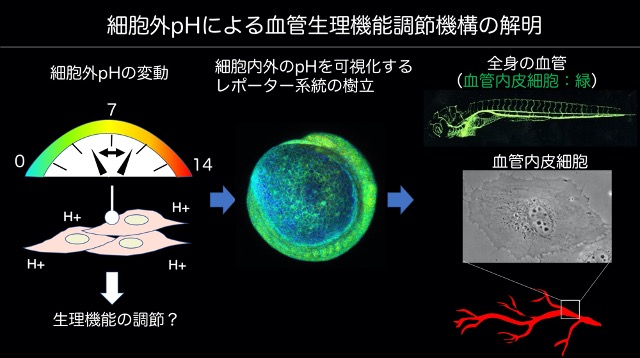
従って、如何にして健康な血管内皮細胞を維持していくかがわれわれの血管が障害から免れることの重要なテーマとなる。我々はこれまで、Vascular endothelial cadherin (VE-cadherin)に注目して血管内皮細胞間接着維持機構を解明してきた。今後もVE-cadherinだけではなく、血管内皮細胞のquiescenceとstabilizationをkey wordsに研究を進めていく。
血管内皮細胞だけでなく、血管内皮細胞を支持する壁細胞(血管平滑筋細胞・周細胞)の発生における役割についても検討している。
OSTNはNa利尿ペプチドファミリー(NP)分子(ANP, BNP, CNP)と一部アミノ酸の相同配列を持ち、Na利尿ペプチドのクリアランス受容体NPR3に結合することを明らかにした。このためにOSTNがNPR3に結合することでNPファミリー分子のクリアランスを阻害して、NPの生理作用を増強することがわかった。
本研究はAMED-CRESTの研究として2013年から開始した。
※画像をクリックすると拡大します







