ファイトプラズマ研究
ファイトプラズマによるユニークな病徴
ファイトプラズマはイネなどの作物を萎縮し枯らすほか、てんぐ巣病や葉化病など1000種類以上の植物に病気を引き起こし、収穫を皆無にするなど、農業に甚大な被害を与える微小な植物病原細菌です。特に、小振りで枝を密生するポインセチアは、全て育種後にファイトプラズマを感染させて出来たものであり、小振りでなければ商品価値はありません。また、花を葉に変える葉化症状を呈する植物は、緑花アジサイのように病気と判明するまでは魅力的で付加価値の高い品種として市場で珍重され高値で取引されるなど、多くの関心を集めてきました。ファイトプラズマにより植物に引き起こされる症状をまとめると次の様になります。
黄 化:葉に生ずる養分欠乏のような黄化症状
萎 縮:茎や葉の生長が害され、著しく萎縮・矮性となる症状
叢 生:側枝が異常に多く生ずる症状
てんぐ巣:側芽が異常に発生し、小枝や小葉が多数密生する症状
葉 化:花弁やがく・雌しべ・雄しべが葉に置き換わる症状
緑 化:葉緑素が分化し花弁などが緑色を帯びる症状
ファイトプラズマ研究の背景
わが国において、ファイトプラズマ病は少なくとも江戸時代にはその発生が記録され、養蚕に必要不可欠なクワでは萎縮病による甚大な被害が続いていました。養蚕は明治時代の最重要産業の一つであったため、1897年の帝国議会決議により調査委員会が設置され6年にわたる原因調査がなされましたが、過度な摘葉による生理障害と結論され、真の原因解明には70年もの年月を要しました。1967年、土居らは電子顕微鏡を用いて、クワ萎縮病等の多くの発病植物の篩部細胞に微小細菌が局在することを世界で初めて発見し、テトラサイクリンにより治癒することから動物マイコプラズマに似た微生物MLOと命名しました。その後世界中でMLOの存在が追認され、少なくとも1,000種類以上の植物に感染し、食料生産確保・環境保全の両面において世界中で甚大な被害をもたらすことが明らかになりました。MLOはヨーロッパのブドウやリンゴ、東アジアのナツメなどの果樹生産において大きな脅威になっており、赤道地帯のココヤシや東南アジア一帯のイネやサツマイモ、キャッサバ、サトウキビなど、開発途上国の農業生産に壊滅的被害をもたらす事例も知られています。
この新たな病原体の一群の発見は、農学のみならず、生物学の新ジャンルを拓く成果として世界中の注目を集めました。しかし、それから約半世紀を経ても培養法や形質転換系は確立されておらず、遺伝子レベルでの研究は遅れており、MLOの実態は長年にわたり不明でした。
MLOからファイトプラズマへ─ファイトプラズマ分子分類体系の確立
MLOの生物分類学的位置づけは長年不明であった上、MLO同士の関係性についても、形態が類似し培養もできないことから、比較解析は困難でした。そのため、ファイトプラズマ病の発生が確認されると、宿主植物の種類ごとにファイトプラズマの名称(系統名)が与えられていました。その結果、国内で約40種類、世界で1,000種類以上ものファイトプラズマ系統が報告される状況になり、MLOの明確な整理・分類の基準が必要とされていました。
こうした状況の中、当研究室においてファイトプラズマの分子レベルでの研究が開始されました(Namba et al., 1993a, b)。当時、分子生物学的な手法による系統分類法が考案されはじめており、当研究室では世界各地に発生するファイトプラズマの16S rRNA遺伝子のPCR増幅系を確立し分子系統解析をおこないました。その結果、MLOはMollicutes (モリキューテス)綱に分類される、分子系統学的に独立した一つの集団であることが明らかになりました(図1)。
従来MLOと称されてきた本微生物が動物マイコプラズマとは独立した集団であることが示されたため、MLOというそれまでの名称をファイトプラズマに変 え、属名をPhytoplasmaとすること、分子系統解析により分類されたサブグループについて、ラテン二名法により暫定種を命名すること等が提案さ れ、承認されました (Namba et al., 1993b)。その後、暫定種の登録が進められ(Sawayanagi et al., 1999; Jung et al., 2002, 2003a, 2003b)、現在では国内で9種、世界で40種程度に整理が進んでいます (Maejima et al., 2014a)。

ファイトプラズマの全ゲノム解読
分類基準の制定により、ファイトプラズマの分子レベルでの研究が世界中で展開されることとなりました。ファイトプラズマの性状解明の手がかりをつかむために次の目標となったのはゲノム解析でした。世界各国でゲノムプロジェクトが進行するなか、2004年、当研究室において、世界で初めてファイトプラズマの全ゲノム解読に成功しました(図2)。この成果は、日本において植物病原細菌の全ゲノムを決定した事例としても初めてのことでした。ファイトプラズマゲノムは、DNA複製や転写、翻訳に必要な基本的遺伝子を保持する一方で、アミノ酸合成系、脂肪酸合成系、TCA回路、酸化的リン酸化に関与する遺伝子を欠き、さらにペントースリン酸回路やPTS、ATP合成酵素に関する遺伝子も欠いていて、自律増殖する生物において最少遺伝子を持つとされるマイコプラズマより代謝関連遺伝子が少ないことが明らかになりました。これはファイトプラズマがマイコプラズマと異なり、細胞内寄生で栄養豊富な植物篩部に生息するため、退行的進化により遺伝子の多くを失ったためと考えられました(図3)。逆に、ファイトプラズマゲノムには、マイコプラズマには無い膜輸送系遺伝子が多数コードされていました(図3)。また、アミノ酸や糖、リンゴ酸などのほか、マンガンや亜鉛、マグネシウム、コバルトなど金属イオンの取り込みに関与する膜輸送系も数多くコードされ、植物の生育に必要なこれらの金属イオンが、ファイトプラズマの感染により宿主から収奪され、ファイトプラズマ病に特徴的な病徴のひとつである養分欠乏に似た症状を引き起こす一因になっている可能性が考えられました(Oshima et al., 2004 Nature Genetics)。本成果により、ファイトプラズマ研究のステージは一気にポストゲノム研究へと押し上げられることとなりました。
 図2 ファイトプラズマの全ゲノムマップ
図2 ファイトプラズマの全ゲノムマップ  図3 ファイトプラズマの代謝系
図3 ファイトプラズマの代謝系 媒介昆虫を決定する分子メカニズム
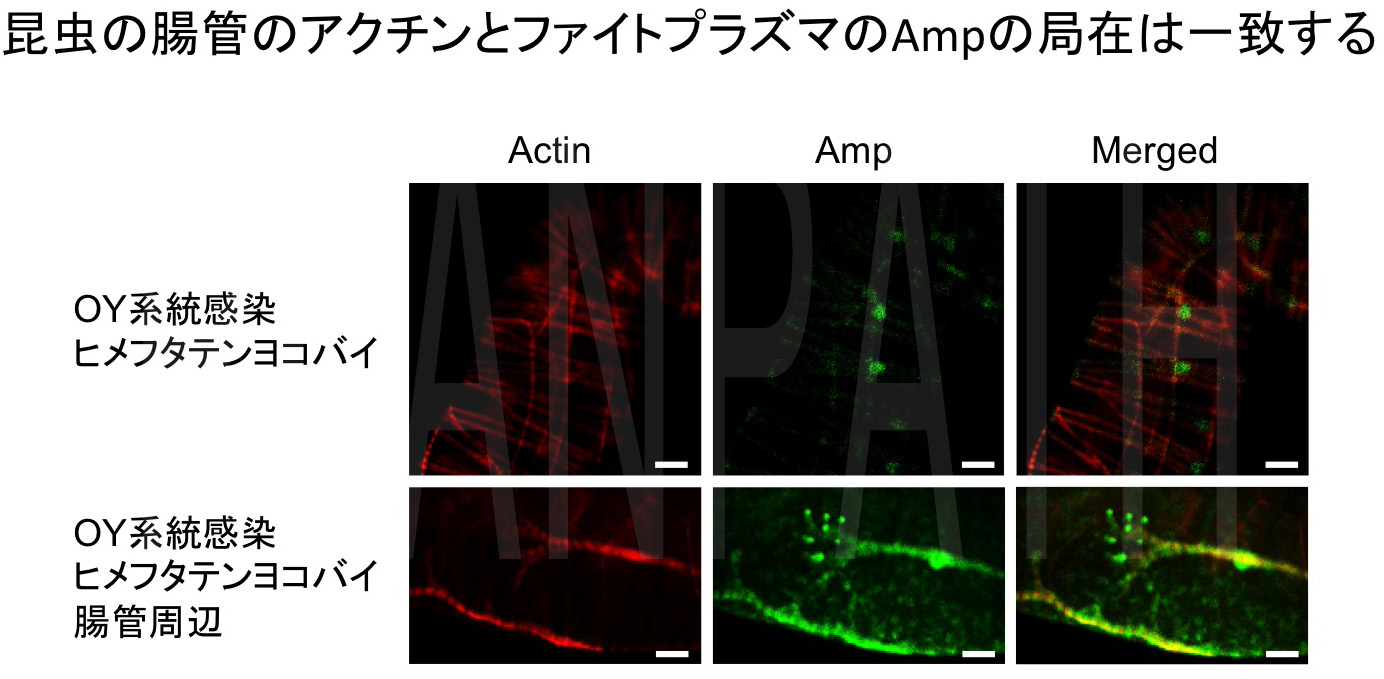
ファイトプラズマは、系統ごとに特定のヨコバイにより媒介され、他のヨコバイにより媒介されることはありません。人の病原体であるマラリアを含め、多くの昆虫媒介性病原体は特定の昆虫によって媒介されますが、その特異性を決めているメカニズムはこれまで不明でした。そこで、ファイトプラズマの媒介昆虫特異性のメカニズムを解明する目的で、まずファイトプラズマの昆虫体内における局在を組織・細胞レベルで解析しました。ファイトプラズマの細胞膜上に豊富に存在するAmpタンパク質に対する抗体を用いて免疫組織化学的観察を行ったところ、ファイトプラズマと昆虫のアクチンとの局在が一致していました(図4)。そこで、Ampを結合させたアフィニティーカラムを作製し、これを用いてAmpと相互作用する昆虫宿主タンパク質を同定したところ、Ampは媒介昆虫の細胞骨格であるマイクロフィラメントを構成するアクチンやミオシンと複合体を形成することが明らかになりました(図5)。さらに、Amp–マイクロフィラメント複合体形成について異なる昆虫を用いて解析を行ったところ、ファイトプラズマ媒介能のある昆虫では複合体の形成が観察されましたが、ファイトプラズマを媒介しない昆虫では複合体の形成が認められませんでした(図6)。これらの結果から、Amp-マイクロフィラメント複合体形成がファイトプラズマの媒介昆虫決定に大きく関与することが明らかになりましたSuzuki et al., 2006 PNAS)。

 図4 昆虫のアクチンとファイトプラズマAmpの局在
図4 昆虫のアクチンとファイトプラズマAmpの局在
 図5 Ampはマイクロフィラメントと複合体を形成する
図5 Ampはマイクロフィラメントと複合体を形成する
 図6 昆虫宿主範囲とAmp-マイクロフィラメント複合体との関連性
図6 昆虫宿主範囲とAmp-マイクロフィラメント複合体との関連性
さらに私達は、全ゲノム解読をおこなったOY-M系統をもとに、昆虫による媒介能を喪失させたファイトプラズマ系統OY-NIM(non-insect-transmissibility)を作出し(Oshima et al., 2001)、異なる切り口からも昆虫媒介のメカニズムの解析を進めました。OY-M系統とOY-NIM系統の遺伝子情報を比較したところ、興味深いことに、OY-NIM系統のゲノム上の配列には変異が見出されなかった一方で、プラスミド上にコードされる膜タンパク質ORF3が失われていることが明らかになりました(図7)(Nishigawa et al., 2002; Ishii et al., 2009a, b)。この結果から、Ampに加えてORF3も、昆虫媒介性において重要な役割を持つ膜タンパク質であると考えられました。
 図7 昆虫媒介能喪失株における膜タンパク質ORF3の欠失
図7 昆虫媒介能喪失株における膜タンパク質ORF3の欠失
私達はさらに、宿主への接着に関わると考えられる膜タンパク質の解析を行いました。多くの病原細菌は宿主細胞への接着・侵入の際に,菌体表面の接着因子が重要な役割を果たします。接着因子は多くの病原細菌で見つかっていますが、ファイトプラズマにおいては見つかっていませんでした。そこで、ファイトプラズマに近縁なマイコプラズマなどの接着因子にみられる、配列保存性の高いアミノ酸モチーフMollicutes adhesin motif(MAM)に着目し、MAMの配列情報を手がかりにファイトプラズマの接着因子ホモログを探索しました。その結果、MAM領域を持つ膜タンパク質P38を見つけました。P38は、ファイトプラズマの宿主である植物や昆虫粗汁液に対して接着能を示し、特に昆虫への接着能にはMAMモチーフが重要であることが明らかになりました(Neriya et al., 2014 FEMS Microbiology letters)。本研究により、これまでマイコプラズマなど近縁の微生物においても不明であったMAMの機能が初めて明らかになりました。本研究を契機として、宿主への接着メカニズムの詳細解明が期待されます。
植物にてんぐ巣症状を誘導する病原性低分子ペプチド「TENGU」
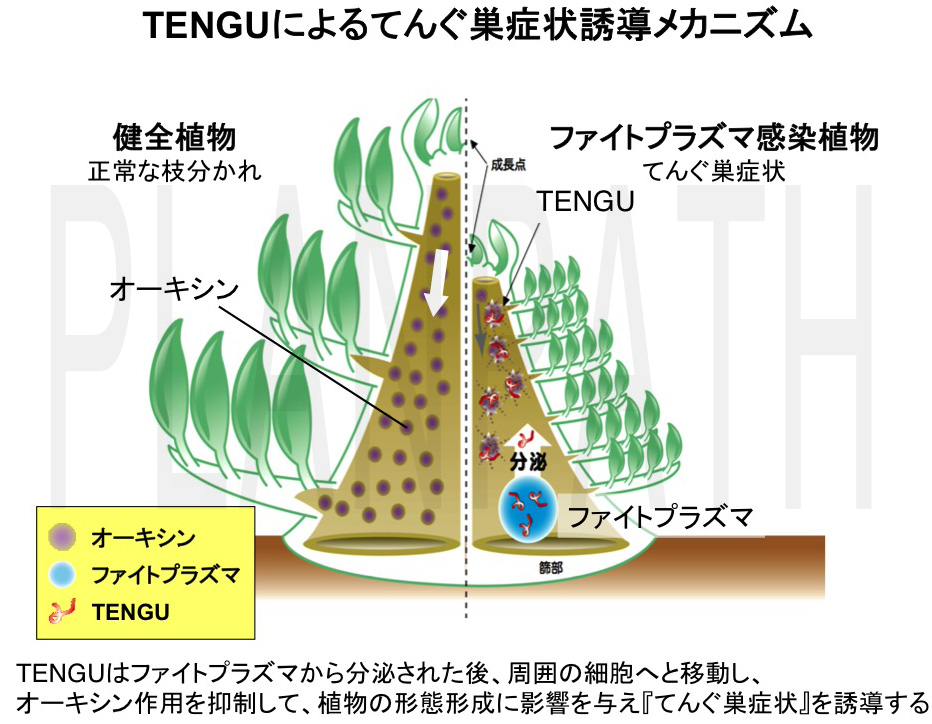
てんぐ巣や葉化などのユニークな病徴は、ファイトプラズマ研究において最も興味深いテーマの一つです。ファイトプラズマが菌体の外へと分泌するタンパク質は、宿主細胞内に直接放出され、宿主細胞の代謝系に直接的に働きかけることができるため、病徴を誘導する病原性因子の最有力候補と考えられました。そこで、ファイトプラズマゲノムにコードされる分泌タンパク質遺伝子に焦点をあて、その病徴誘導活性のスクリーニングをおこないました。その結果、分泌タンパク質のうちの一つを発現させた植物において、草丈の伸長が抑えられる「萎縮症状」と、枝分かれが異常に増加する「叢生症状」が認められ、これら症状はファイトプラズマ感染植物と酷似していました(図8)。 萎縮・叢生症状を呈する植物は、日本では古来より「てんぐ巣病」と呼ばれていたことから、萎縮および叢生症状を誘導するこの分泌タンパク質をてんぐ巣病の誘導因子「TENGU」と命名しました。TENGUはわずか38アミノ酸からなるタンパク質で、植物に形態変化を誘導する植物病原細菌由来のペプチド性因子は、世界で初めての発見でした。ファイトプラズマは篩部局在性ですが、ファイトプラズマ感染植物組織におけるTENGUの動態を調べたところ、TENGUは植物の茎頂分裂組織(成長点)や側芽の分枝領域の細胞にまで移行していることが分かりました。また、TENGUを発現する形質転換植物において特異的に発現が変動する植物遺伝子群を、マイクロアレイ解析によって特定した結果、植物ホルモンの一つであるオーキシンに関連した遺伝子群の発現レベルが大きく低下していました。以上の結果より、TENGUはファイトプラズマから分泌された後、周囲の細胞や茎頂分裂組織へと移行し、オーキシン関連経路を抑制して植物の形態形成に影響を与えることにより、てんぐ巣症状を引き起こすことが示唆されました(図9)(Hoshi et al., 2009 PNAS)。
 図8 ファイトプラズマ病原性因子「TENGU」の同定
図8 ファイトプラズマ病原性因子「TENGU」の同定
 図9 TENGUによるてんぐ巣症状誘導メカニズム
図9 TENGUによるてんぐ巣症状誘導メカニズム
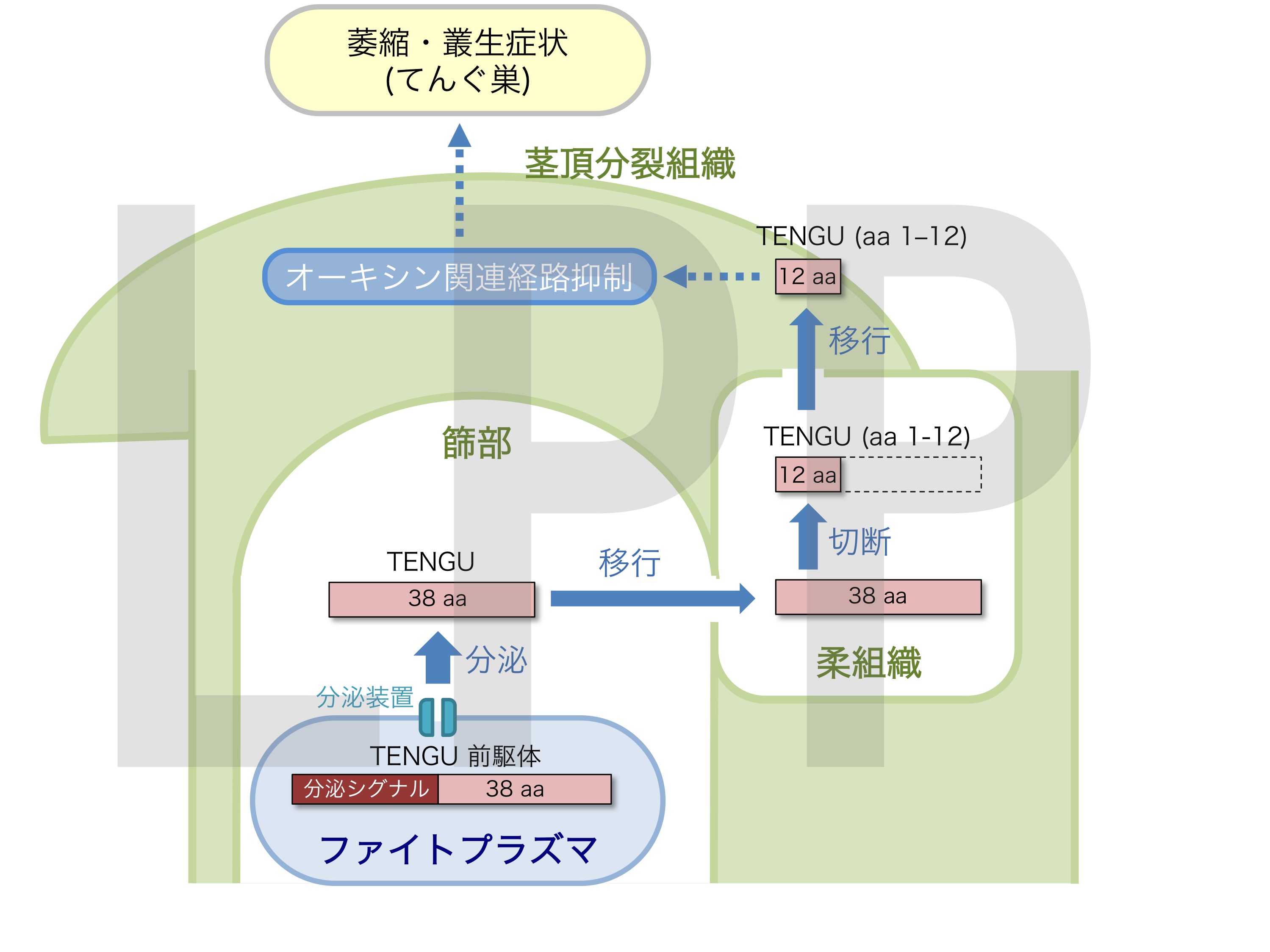
さらに私達はTENGUの病徴誘導領域を絞り込み、全長38アミノ酸のうちN末端側のわずか11アミノ酸がてんぐ巣症状を引き起こす活性を持つことを明らかにしました(図10)。そこで、TENGUが植物内で切断される可能性を検証したところ、実際にin vitro と in planta の両条件下で、TENGUが切断されることが分かりました。ペプチドシーケンス及びTOF-MSを用いた解析から切断部位の特定をした結果、N末端側は病徴誘導領域である11アミノ酸を含む長い断片として生成されることが明らかになりました。また、TENGUの切断はセリンプロテアーゼ阻害剤であるPMSFにより阻害されたため、植物が持つセリンプロテアーゼによりTENGUが切断されていると考えられました。加えて、TENGUのファイトプラズマ間における保存性を解析したところ、主要グループの1つであるAYグループにおいてTENGUの配列や機能、被切断特性が広く保存されていることが明らかになりました。以上の結果から、TENGUは植物内で切断され、そのN末端側のペプチドがてんぐ巣症状を誘導すること、さらにTENGUによるてんぐ巣症状の誘導機構が複数のファイトプラズマに保存されていることが示唆されました(図11)(Sugawara et al., 2013 Plant Physiology)。
 図10 TENGUの病徴誘導活性を持つ領域の特定
図10 TENGUの病徴誘導活性を持つ領域の特定
 図11 TENGUによる萎縮・叢生症状誘導モデル
図11 TENGUによる萎縮・叢生症状誘導モデル
一方、ファイトプラズマに感染した植物は、しばしば種子を作れず子孫を残せない「不稔症状」を伴った病徴を引き起こします。不稔症状は、果実や種子の生産に致命的な被害を与える重大な病気の一つですが、その原因物質やメカニズムについてはこれまで不明でした。当研究室では、不稔症状を引き起こす病原性因子が「TENGU」であり、一つの病原性因子が「天狗巣」と「不稔」という2つの異なる症状を引き起こす、非常に興味深い現象を突き止めました。植物にTENGUを発現させると、花粉が奇形となり、雄性不稔症状が起こることを発見しました(図12)。TENGUを発現させた植物の花芽では、花の稔性に関わる複数の植物ホルモン(ジャスモン酸およびオーキシン)の蓄積量が低下することが明らかになりました。さらに、ジャスモン酸の発現に必要な2つの遺伝子、AUXIN RESPONSE FACTOR 6 (ARF6) およびARF8 の発現量が、TENGU発現植物およびファイトプラズマ感染植物いずれにおいても抑制されることを明らかにしました。従って、ファイトプラズマに感染した植物では、TENGUの作用により植物のARF6およびARF8の発現が抑制され、植物ホルモン量が低下し、不稔症状を引き起こすと考えられました。このように「TENGU」は2つの異なる植物ホルモンに作用するという、病原性因子としてこれまでに例のないユニークな機能を持つことが明らかになりました(Minato et al., 2015 Scientific Reports)。
 図12 TENGU発現植物における花粉の成熟不全と不稔
図12 TENGU発現植物における花粉の成熟不全と不稔 ファイトプラズマが花を葉に変えるメカニズム
花が葉になる「葉化(phyllody:phyllo-(葉)+-ody(になる変化))」や、花から若芽が出現する「つき抜け」などの花器官における病徴も、ファイトプラズマが引き起こす非常にユニークな病徴です。近年、花の形態形成に関わる植物の遺伝子群が明らかになってきたため、これらの中でも特に各花器官(がく、花弁、雄蕊、雌蕊)の形成を制御するA、B、C、D、E遺伝子に着目し、ファイトプラズマの感染による影響を解析しました。A~Eにより表されるこれらの遺伝子はいずれもMADSドメイン転写因子をコードしており、それらがヘテロ四量体を形成する際の組合せにより、各花器官への分化が決定されます(カルテットモデル)。葉化症状が見られた花器官では、そのヘテロ四量体の形成に必要な遺伝子の一部の発現量が有意に減少していることが判明しました(図13)。興味深いことに、発現量が減少した遺伝子の種類に一貫性はなく、例えば葉化したがくでは、がくの形成に必要とされるA、E遺伝子のうち、A遺伝子の発現が抑制されていた一方、同じく葉化の見られた花弁では、花弁の形成に必要とされるA、B、E遺伝子のうち、A遺伝子ではなくB遺伝子の発現が抑制されていました。以上により、ファイトプラズマが花の形成に関わる遺伝子群に関して花器官ごとに巧みに発現制御をおこなうことによって、葉化を引き起こしていることが示されました(Himeno et al., 2011 Plant Journal)。

本研究は分子レベルで葉化症状の原因を説明づけた世界で初めての成果となりました。今回得られた成果は、植物の花器官形成メカニズムの解明にもつながる普遍性があり、園芸品種の育成に応用することで病原体フリーの葉化品種の開発も期待されています。

図13 花が形成される際の植物宿主側の遺伝子変動
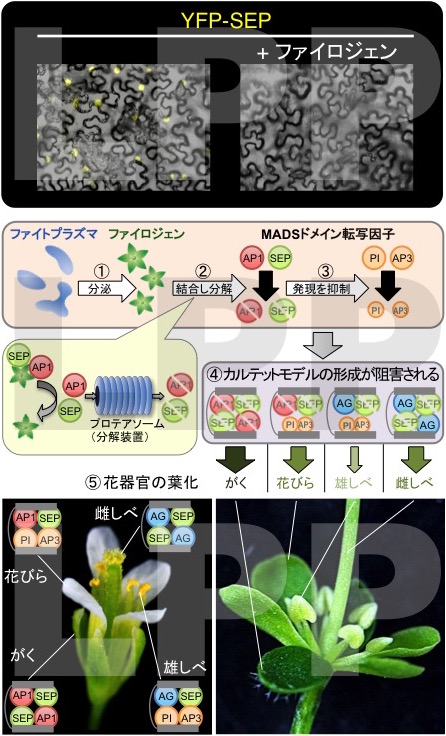
さらに、花が葉になる葉化症状の原因遺伝子が、数多くのファイトプラズマのゲノムに共通して存在する遺伝子群であることを発見し「ファイロジェン(phyllody-inducing gene family(葉化誘導遺伝子ファミリー)の略)」と命名しました。ファイロジェンは、わずか91アミノ酸からなる分泌タンパク質をコードしており、植物において発現させると、ファイトプラズマによる症状と同様に花が葉化しました(Plant J cover)。ファイロジェンの働きを調べたところ、まずファイロジェンはカルテットモデルを構成するMADSドメイン転写因子群のうち少なくともA、Eの転写因子に結合する性質を持っていることが分かりました。そこで、実際にA、E転写因子をファイロジェンとともに植物細胞に導入したところ、それら転写因子が分解されることが明らかになりました。また、タンパク質分解装置(プロテアソーム)を阻害するようにあらかじめ処理すると転写因子の分解が阻害されたため、ファイロジェンが植物のプロテアソームを利用してMADSドメイン転写因子群を分解していることが確かめられました。また、A、E転写因子はB転写因子の発現を誘導することが知られていますが、ファイロジェンを発現させた植物では逆にB転写因子の発現が抑制されていました。つまり、A、E転写因子が細胞内で分解されたため、B転写因子の誘導が阻害されたと推定されます(図14)。
以上の結果から、ファイトプラズマに感染しファイロジェンが分泌された植物では、各花器官の形成に必要なカルテットモデルを構成する転写因子の多くが、分解または発現抑制され、これが原因で各花器官の葉化が起こり、花から新たに枝葉が形成されることが説明されます(Maejima et al. 2014b Plant Journal)。

これらの研究は分子レベルで葉化症状の原因を説明づけた世界で初めての成果となりました。今回得られた成果は、植物の花器官形成メカニズムの解明にもつながる普遍性があり、園芸品種の育成に応用することで病原体フリーの葉化品種の開発も期待されています。

図14 ファイロジェンの作用メカニズム 動物・植物間のホストスイッチングに伴う環境適応
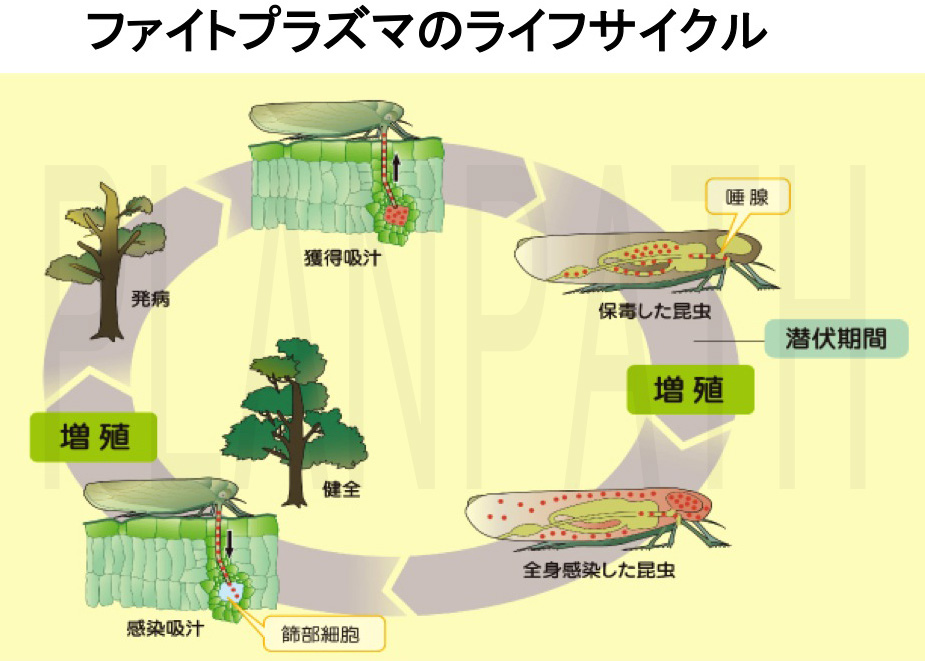
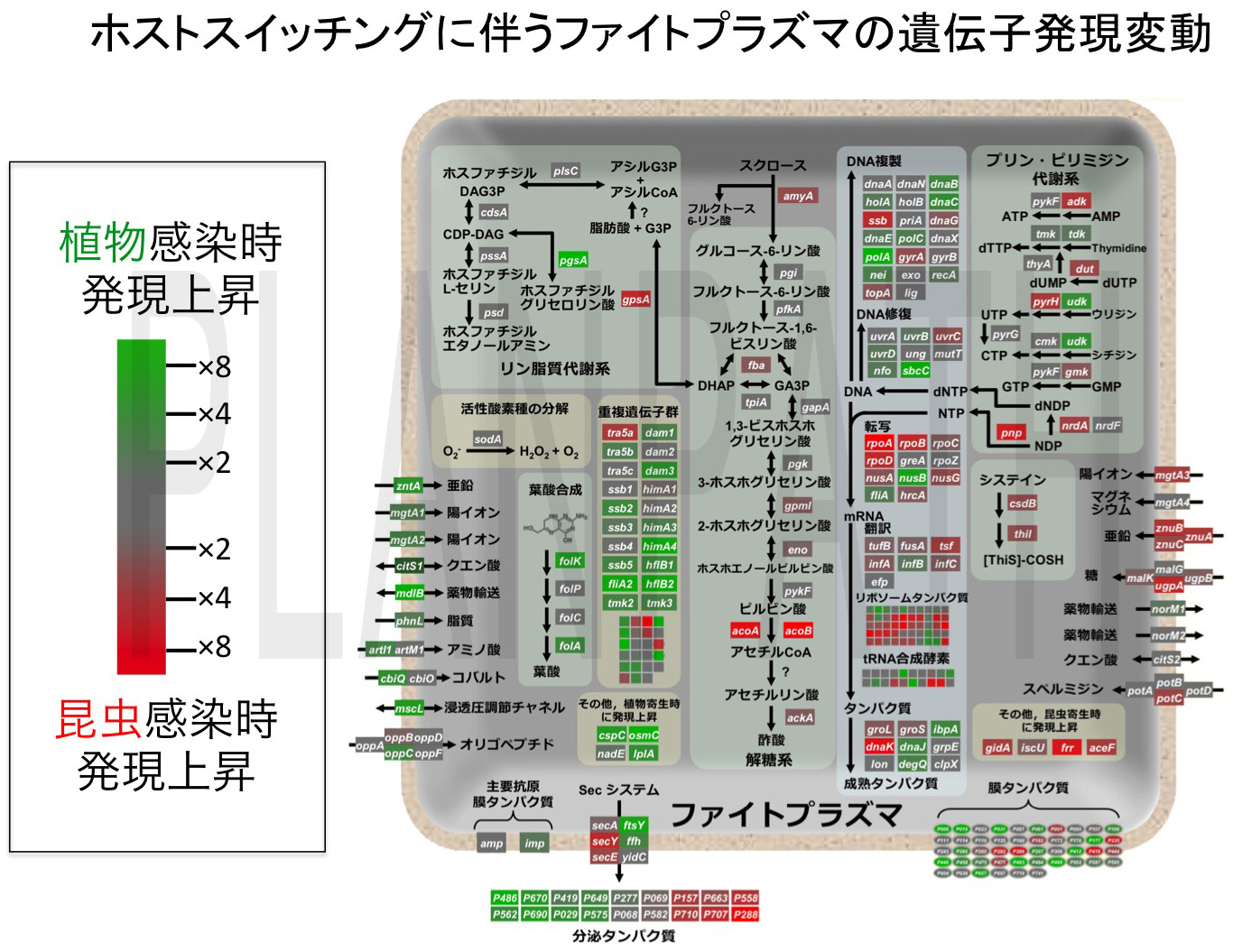
ファイトプラズマは昆虫-植物の宿主間を水平移動する「ホストスイッチング」により感染を拡大します(図15)。昆虫と植物という全く異なる生物界の宿主に寄生する能力には興味が持たれてきましたが、その仕組みはこれまで謎に包まれていました。ホストスイッチングにはファイトプラズマの遺伝子発現の切り替えが伴うと予想されたため、それら遺伝子を網羅的に調べるために、ゲノムデータ(図2)をもとに世界初となるファイトプラズマのDNAマイクロアレイを作製しました。マイクロアレイ解析の結果、ファイトプラズマは植物宿主と昆虫宿主とを移動するたびに、ゲノム全体の約1/3に相当する遺伝子の発現量を切り替えていることが明らかになりました(図16)(Oshima et al., 2011 PLoS One)。特に、ファイトプラズマはそれぞれの宿主に合わせて、物質輸送を行うトランスポーターや浸透圧を調節するチャネル、糖を分解する酵素、宿主細胞内で働く分泌タンパク質などを巧みに使い分けている点に興味が持たれました。そこで、実際に植物感染時に働く浸透圧調節チャネルの機能を、阻害剤を用いて抑制したところ、ファイトプラズマの増殖を部分的に抑えることに成功しました。この結果は、ホストスイッチングの阻害・抑制が、特効薬が無く防除や予防が困難なファイトプラズマ病の新規防除法につながる可能性を示す成果です。
 図15 ファイトプラズマのライフサイクル
図15 ファイトプラズマのライフサイクル
 図16 ホストスイッチングに伴うファイトプラズマの発現変動
図16 ホストスイッチングに伴うファイトプラズマの発現変動
このように、ファイトプラズマはホストスイッチングに伴いダイナミックに遺伝子の発現を切り替えていることが明らかとなったため、次にファイトプラズマの遺伝子発現制御機構について研究を進めました。多くの細菌は、周囲の環境変化に応じてσ(シグマ)因子と呼ばれる転写因子を使い分けることで遺伝子発現を制御しています。ファイトプラズマはRpoDとFliAという2種類のσ因子を持ちますが、中でもRpoDはファイトプラズマ間で保存性が高い上に昆虫体内で特に発現量が上昇するため(Oshima et al., 2011 PLoS One)、ホストスイッチングに関与する可能性が考えられました。そこで、in vitro でのRNA転写系を構築し、ファイトプラズマのRpoDにより発現制御される遺伝子を解析しました。その結果、ハウスキーピングな遺伝子に加えて、病原性や宿主相互作用に関わる様々な遺伝子の発現を制御していることが明らかになりました(Miura et al., 2015 Scientific Reports)。また、他の全ゲノム解読されたファイトプラズマにおいてもRpoDに同様の活性があることを、in silico 解析により明らかにしました。RpoDは一般に、恒常的に発現してハウスキーピング遺伝子の発現を司るσ因子として知られていましたが、本研究により、ファイトプラズマのRpoDは他の細菌よりも複雑な機能を担うことが示唆されました。これは、ゲノムの退行的進化により獲得された遺伝子発現制御機構であると考えられ、ファイトプラズマのホストスイッチング機構の解明に繋がる重要な成果であると考えられます。
ファイトプラズマ研究のこれから
ファイトプラズマ研究に分子のメスが入ってから20年、全ゲノム解読から10年が経過しました。この間、ファイトプラズマ研究は目覚ましい勢いで進展し、当研究室の研究チームは常に世界をリードし続けてきました。特にこの10年のポストゲノム研究により、当初は想像もできなかったようなファイトプラズマの実体が私たちによって次々と解明されてきました。しかし、明らかになった部分はまだほんの一部であり、依然としてその性状の多くは謎に包まれています。例えば、ファイトプラズマの極限にまで切り詰められてもなお生命として存在する代謝系の動植物宿主との物質のやり取りを通じた動的解明はまだ終わっていません。また、ファイトプラズマが植物に引き起こすユニークな病徴の生物学的意義や植物・ファイトプラズマの共進化の意義、植物宿主域の決定メカニズムの詳細も未だ完全には解明されていません。そして、もちろん宿主である植物・昆虫のファイトプラズマ感染に対する防御応答や共生のメカニズム解明へと私たちの研究の展開にはボーダーはありません。しかし、ゴールはもう間近に迫っています。ファイトプラズマ研究はこの20年の間に驚異的に進化しましたが、これまでの知見を基盤に、新たなパラダイムを構築することが、今後の研究展開の重要な鍵となります。世界の食料生産・森林資源・農地・緑地・住地環境に甚大な被害と脅威を与え続けているファイトプラズマ病の治療・防除・予防技術のさらなる進化に向けて、今後の研究の飛躍が期待されます。